




-
开放科学(资源服务)标识码(OSID):

-
我国是苹果生产大国,栽培面积和产量都居世界之首. 苹果绵蚜(Eriosoma lanigerum (Hausmann))属于半翅目(Hemiptera)瘿绵蚜科(Pemphigidae)绵蚜属,又叫血色蚜虫、赤蚜、绵蚜等,原产于北美洲东部,随苗木传播至70多个国家和地区[1]. 20世纪初苹果绵蚜传入我国山东威海,从20世纪末开始,入侵范围不断扩大,危害加重,先后传到河北、河南、甘肃、天津、山西、新疆等地,对北方苹果产业造成巨大危害[2]. 苹果绵蚜主要刺吸危害苹果树的嫩枝、主干,造成营养失衡,影响开花结果,严重情况下会造成果树死亡.
已有研究发现,目前苹果生产中的主栽品种“红富士”的父本“小国光”及与其母本“红元帅”亲缘关系较近的“新红星”品种相对抗蚜,而“红富士”高感苹果绵蚜[3-4]. 在果树生长期调查苹果绵蚜在不同苹果品种上的数量,发现“红富士”的绵蚜数量(137.7~172.9头/株)显著高于“乔纳金”(20.0~46.1头/株)与“新红星”(23.3~41.9头/株);在苹果树落果后调查苹果绵蚜对不同苹果品种的危害率及虫落数量,发现苹果绵蚜对“红富士”的危害率为64.8%,显著高于“青香蕉”“乔纳金”“新红星”“小国光”的危害率(26.6~38.5%). 尹学伟等[5]对“红富士”“金帅”“昭锦108”接种苹果绵蚜,将苹果绵蚜危害苹果树后产生的肿瘤进行石蜡切片研究发现,“红富士”是感性品种,对苹果绵蚜抗性最弱,“昭锦108”抗性最强. 通过模拟田间温度发现,苹果绵蚜取食“红富士”的发育速度、繁殖数量以及内禀自然增长率、净增值率、周限增长率均显著高于“新红星”“小国光”[6],这与新西兰研究结果类似[7-8]. 通过EPG刺吸电位(Electrical Penetration Graph,EPG)技术进一步研究发现,苹果绵蚜取食“新红星”和“小国光”韧皮部阶段产生的E1波(取食)总时间显著长于“红富士”,而E2波(分泌唾液)总时间显著短于“红富士”[9]. 由此可见,“红富士”“珊夏”“嘎啦”是苹果绵蚜的高感品种,“金冠”“首红”“乔纳金”“小国光”“昭锦108”“新红星”等则是高抗品种,这可能是苹果绵蚜危害加重的重要原因之一.
在植物中,12-氧-植物二烯酸还原酶(12-oxo-phytodienoic acid reductase,OPR)属于古老黄酶(Old Yellow Enzyme,OYE)家族,是一种黄素单核苷酸(Flavin Mononucleotide,FMN)依赖的氧化还原酶,典型的OPR蛋白只有1个Oxidored_FMN结构域. 植物OPR家族成员表现出多种生物学功能,可以被茉莉酸(Jasmonic Acid,JA)、乙烯(Ethylene,ET)、脱落酸(Abscisic Acid,ABA)和水杨酸(Salicylic Acid,SA)等各种激素以及各种生物和非生物胁迫诱导[10].
OPR是亚麻酸合成茉莉酸途径中的一个关键酶,茉莉酸及其衍生物统称为茉莉素(Jasmonates,JAs),是广泛存在于植物中的一类植物内源激素,对植物生长和发育具有广泛的生理效应,同时可作为内源信号分子参与植物的抗逆反应. 植物遭遇昆虫取食或机械损伤后,激发茉莉酸的生物合成过程[11],JA生物合成始于细胞膜释放的α-亚麻酸,接着在叶绿体中,α-亚麻酸在脂氧合酶(lipoxygenase,LOX)、丙二烯氧化合酶(Allene Oxide Synthase,AOS)、丙二烯氧化环化酶(Allene Oxide Cyclase,AOC)作用下转化为12-氧-植物二烯酸(12-oxo-phytodienoic acid,OPDA)[12-14],继而在过氧化物酶体中,OPDA被OPR还原为茉莉酸的前体OPC - 8∶0[15].
在高等植物中,第一个被发现的OYE成员为拟南芥中的AtOPR1[16],随后在其他高等植物中也相继发现了OPR基因,其中包括拟南芥的另外两个OPR基因AtOPR2和AtOPR3[17-18],番茄中的3个OPR基因[19-20],豌豆中的6个OPR基因[21],玉米中的8个OPR基因[22]以及水稻中的13个OPR基因[23-25]. OPR在茉莉酸的合成过程中,是控制JA合成的最后步骤,即催化OPDA生成JA的前体OPC - 8∶0[26-28]. OPDA有4种立体异构体的结构,根据底物偏好性将OPR分为2类,OPR Ⅰ主要催化9R,13R-OPDA,也催化9S,13R-OPDA,产生一些副产物,几乎不催化另外2种13S构型的OPDA;OPRII能有效地利用OPDA的4种异构体,是唯一能够将天然的9S,13S-OPDA转化为相应的9S,13S - OPC - 8∶0,即(+)-7-epi-JA的前体物质,因此只有OPRII亚类基因参与内源茉莉酸的合成[29].
本研究根据项目组前期完成的不同苹果品种被苹果绵蚜危害后的转录组学和蛋白质组学,鉴定了茉莉酸代谢通路中OPR基因,并通过生物信息学和qPCR分析了茉莉酸代谢通路中对苹果绵蚜起抗性作用的OPR基因,旨在为阐明抗性品种对苹果绵蚜的抗性机理奠定基础.
HTML
-
2019年3月选择烟台栖霞苗圃1年生苹果品种“红富士”(RF)、“新红星”(SM)、“小国光”(RG)各50棵. 苹果绵蚜采集于山东烟台,带回实验室用苹果幼苗饲养.
-
5—6月选取1年生“红富士”(RF)、“新红星”(SM)、“小国光”(RG)苹果苗,选定接种的枝条后,在距顶芽8~10 cm处接种25头蚜虫. 将苹果树放到培养箱中,条件为:14 h光照,10 h黑暗,温度23 ℃,相对湿度60%. 分别在12 h、5 d和18 d取样,用灭菌的毛笔清除蚜虫、虫蜕等残余污染物,用灭菌的剪刀去掉叶片,剪下苹果绵蚜危害处的6 cm左右的枝条,装到无酶离心管中,液氮速冻,以未接种蚜虫的苗木为对照,每个处理重复3次. 由安诺优达生物科技有限公司对0 h、危害12 h、危害5 d、危害18 d的4个时间段样本进行转录组测序,对0 h、危害12 h、危害5 d后的3个时间段样本进行蛋白组测序.
-
使用TBtools做基因表达量热图;使用NCBI(
https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.Cgi )、Smart(http://smart.embl.de/smart/save_user_preferences.pl )对基因功能域进行预测,使用DOG 2.0进行做图;蛋白质多序列比对通过Clustalw完成(https://www.genome.jp/tools-bin/ ),蛋白质理化性质分析通过Ex-pasy(https://web.expasy.org/protparam/ )进行;使用MEGA5软件构建系统进化树. 采用SPSS软件对试验数据进行独立样本T检验. -
使用Primer 5.0软件根据目的基因MdOPR1,MdOPR4的序列设计引物,设计的引物序列见表 1.
-
以EF-α为内参基因,按以下体系加样(表 2).
-
上机前预扫:将靶标蛋白列表导入SpectroDive软件,匹配相应的Library(即上述DDA库),软件会自动计算目标蛋白理论肽段序列的质荷比,并保留符合以下条件的独特肽段:①目标蛋白特有的肽段;②无漏切位点;③无可变修饰;④电荷数:2~3. 导出以上列表,设置质谱方法上机一针mix pre-scan.
PRM方法导出:将预扫的raw data导入SpectroDive软件,根据iRT标准肽段进行保留时间校正,设置scheduled方法,并导出.
PRM模式检测:色谱条件,流动相A相:ACN-H2O-FA(0∶100∶0.1,v∶v∶v);流动相B相:ACN-H2O-FA(80∶20∶0.1,v∶v∶v);流速:300 nL/min;梯度洗脱条件:0~60 min,8%~25% B;60~79 min,25%~45% B;79~80 min,45%~100% B;80~90 min,100%B. 主要扫描参数包括:MS2分辨率30 000,分离窗口(Isolation Window)1.2 m/z,AGC设置为1e5,最大累计时间80 ms,HCD能量28.
1.1. 材料
1.2. 转录组、蛋白组取样
1.3. 生物信息学分析
1.4. 抗虫基因荧光定量PCR引物设计
1.5. qRT-PCR验证
1.6. PRM验证
-
从NCBI查找拟南芥、小麦、玉米和丹参等植物的OPR基因,下载其蛋白序列,通过本地Blast方法,以e-5为标准,在使用的基因库中共找到16个基因,通过功能域分析,11个基因含有FMN结构域,3个基因含有PLN02411结构和FMN结构域,2个基因无FMN结构域,因此,共14个基因属于OPR家族. 将这14个基因与玉米ZmOPR5和水稻OsOPR7的序列进行整理,使用在线多序列比对工具进行比对(图 1),利用NCBI数据库进行分析,共有4类关键位点,其中活性位点15个,其中在OsOPR7蛋白序列中位于第245位的组氨酸(His,H)对还原酶的底物选择起到关键作用. 底物结合位点有6个,包括T34,W109,H186,H189(N189),Y191,R344. FMN结合位点10个,包括P32,T34,G65(A65),Q107,R238,G321,S341(A341),G343,R344,L345. 催化残基只有Y191.
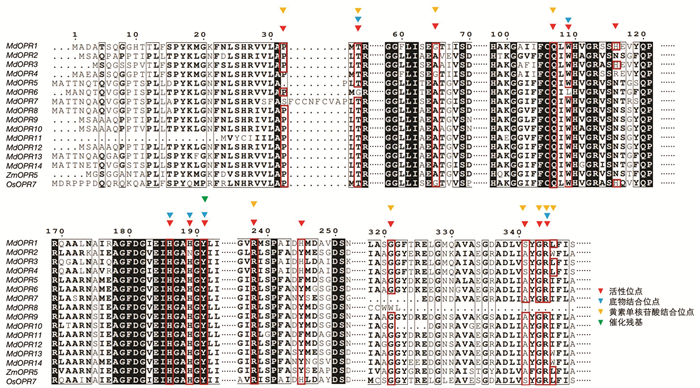
-
利用MEGA5软件,将筛选到的14个基因与拟南芥、水稻、玉米、云杉、莱茵衣藻的OPR基因序列进行系统进化树比对. 根据结果来看,OPR家族分为两大枝,苹果树中的MdOPR1,MdOPR2,MdOPR4共3个基因与水稻OsOPR7、拟南芥AtOPR02-1(AtOPR3)、玉米ZmOPR7,ZmOPR8聚类在OPRII亚类,苹果树另外的11个基因与OPRI亚类聚合在一起. 因此,在苹果树中有3个基因(MdOPR1,MdOPR3和MdOPR4)属于OPRII亚类,可能参与茉莉酸的抗虫反应(图 2).
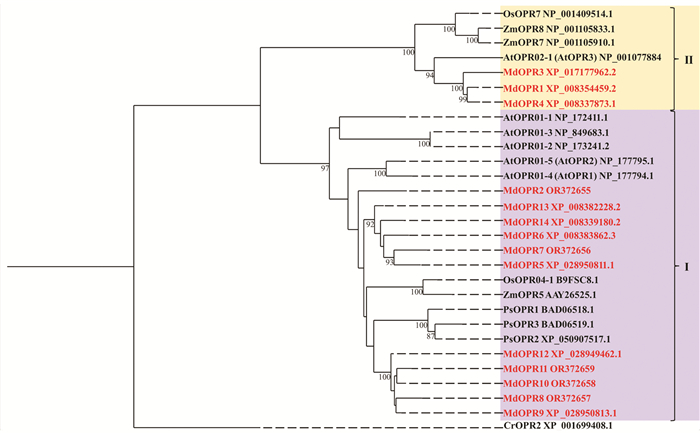
-
通过鉴定分析筛选到的14个OPR基因,将其命名为MdOPR1~14. 利用生物信息学方法对基因编码的蛋白理化性质分析结果如表 3所示,氨基酸序列长度为305~398个氨基酸残基AA. 从等电点来看,聚合在于OPRII亚类的3个基因(MdOPR1,MdOPR3和MdOPR4)等电点均大于7,其余蛋白等电点为5.10~6.80(表 3),也即OPRII亚类的基因等电点均大于归属于OPRI亚类的基因,这也与Schaller等[29]的研究结果相符.
-
对筛选出的14个OPR家族基因进行转录组表达量分析(图 3),其中差异基因有5个(MdOPR2,MdOPR5,MdOPR6,MdOPR12和MdOPR14). 根据转录组分析结果,对于属于OPRII亚类的3个基因,基因MdOPR1表达量在“红富士”和“新红星”中均呈现先升高后下降的趋势,在苹果绵蚜危害12 h时表达量最高;基因MdOPR3表达量在“红富士”和“小国光”中呈现出先升高后下降的趋势,在“新红星”中表达量呈下降趋势;基因MdOPR4在3个品种中,表达量均呈现出逐步升高的趋势. 基因MdOPR5和MdOPR14的表达量在“红富士”和“小国光”中均呈下降趋势.
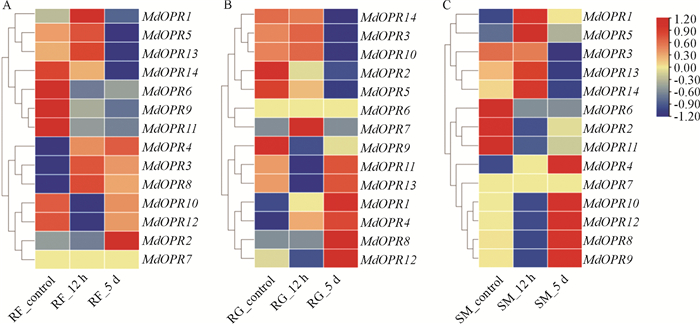
-
将14个OPR基因在蛋白组中进行表达量分析,共筛选到6个(MdOPR1,MdOPR2,MdOPR4,MdOPR10,MdOPR12和MdOPR13)具有显著性差异基因,MdOPR1基因在3个品种中的表达量均呈上升趋势,苹果绵蚜危害5 d时表达量最高;MdOPR4基因在抗性品种“小国光”中表达量呈上升趋势,而在“红富士”和“新红星”品种中表达量呈下降趋势. 基因MdOPR12,MdOPR13基因在“小国光”中表达量呈下降趋势,在“新红星”和“红富士”中表达量先上升后下降,苹果绵蚜危害12 h时表达量最高;MdOPR2基因在“小国光”中表达量呈下降趋势,在“红富士”和“新红星”中表达量总体呈上升趋势;MdOPR10基因在3个品种都呈下降趋势(图 4).
-
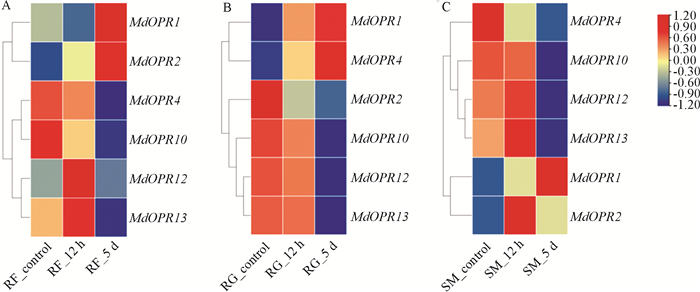
对本研究筛选得到的苹果树MdOPR1和MdOPR4两个聚类在OPRII亚类的基因进行qRT-PCR验证,结果与转录组数据相符. 苹果树在被苹果绵蚜危害后,MdOPR1基因在感性品种“红富士”中表达量先上升,后下降,随后再上升;在抗性品种“小国光”和“新红星”中表达量先上升,后下降,在“红富士”和“新红星”中危害12 h的表达量显著高于危害0 h;在“小国光”中,危害5 d时的表达量最高,显著高于危害0 h. 在相同实验条件下,MdOPR4基因在3个品种中表达量趋势均为先上升,再下降,在感性品种“红富士”中,虽然上升,但是差异不显著,在抗性品种“小国光”和“新红星”中,危害12 h时,表达量显著高于危害0 h,分别在5 d和12 h时表达量到达最高值(图 5).
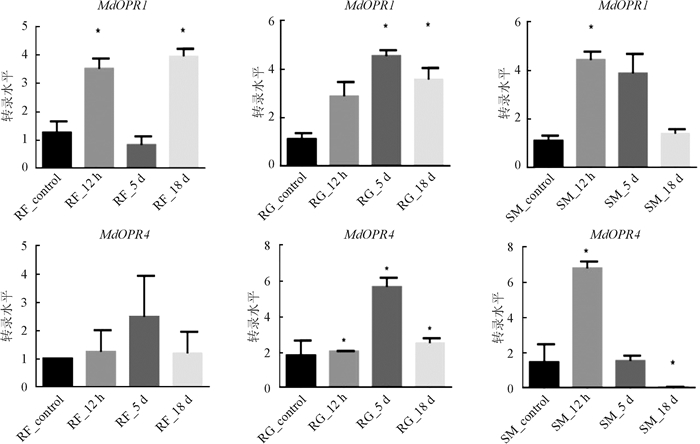
-
对MdOPR1进行PRM验证,结果发现该蛋白在抗性品种“小国光”中上调1.14倍,在“新红星”中上调倍数最多,上调1.51倍,在感性品种“红富士”中上调倍数最低,上调1.13倍,这与蛋白组结果一致,结果显示该蛋白与抗性相关.
2.1. 苹果树OPR基因鉴定
2.2. 系统进化树
2.3. 基因理化性质
2.4. 转录组表达量分析
2.5. 蛋白组表达量分析
2.6. qRT-PCR验证
2.7. PRM验证
-
茉莉酸及其衍生物信号在植物抗虫、抗病方面有重要的作用. 茉莉酸及其衍生物本身对昆虫无毒,但将茉莉酸及其衍生物喷施在植物上却可以诱导植物次生化学物质和系统抗性的产生,避免昆虫对植物的伤害. 茉莉酸及其衍生物可能通过诱导多酚氧化酶(Polyphenol Oxidase,PPO)和蛋白酶抑制剂,造成植食性动物蛋白质营养获取障碍,从而抑制昆虫生长. 在不同的物种中,茉莉酸及其衍生物可诱导植物产生不同的次生代谢物,如尼古丁、芥子油苷、异类黄酮、生物碱、萜类、酚类、醌类、胼胝质、硫代葡萄糖苷、生氰葡萄糖苷、酰基蔗糖等,影响昆虫对营养物质的吸收,或干扰植食动物神经信号传递[30-33]. 玉米中的双突变体opr7opr8的JA水平在所有器官中都显著降低,双突变体导致了大量的发育缺陷和对病原体及昆虫的极端易感性[34]. 因此,本文对苹果树被苹果绵蚜危害的转录组、蛋白组的茉莉酸通路中的OPR基因进行了研究.
“红富士”是以“红元帅”为母本、“小国光”为父本杂交得到,“新红星”是由“元帅”芽变得到,这3个品种具有一定的亲缘关系,而对苹果绵蚜的抗性不同. 因此,本文选择这3个品种苹果树进行抗蚜性研究. 本研究通过本地Blast和功能域预测,共鉴定出14个OPR基因,其进化分析结果表明,MdOPRs可分为两类,MdOPR1,MdOPR3和MdOPR4与其他植物具有抗虫作用基因聚类在一起. 有研究表明,玉米OPR基因受JA调控[20];在水稻的OPR家族中,目前仅有处在OPRII亚类的OsOPR7被报道能够催化OPDA. 通常,进化树中亲缘关系越近,其具有相似基因功能的可能性也越大,因此,推测MdOPR1,MdOPR3和MdOPR4与AtOPR02-1(AtOPR3)有相似功能,可以有效催化9R,13R-OPDA,使其转化为JA的前体OPC-8∶0[29].
本研究对苹果绵蚜危害后的苹果树转录组、蛋白组中的茉莉酸通路进行研究,对筛选到的14个OPR基因,MdOPR1在3个品种中抗性反应一致,随着苹果绵蚜危害时间增加表达量增加;MdOPR4基因在抗性品种“小国光”中随苹果绵蚜危害时间增加表达量增加,在“新红星”和“红富士”中随苹果绵蚜危害时间增加表达量先升高后下降,分别在12 h和5 d后达到最高值;MdOPR6基因在“红富士”中随苹果绵蚜危害时间增加表达量增加;MdOPR2在“红富士”中随苹果绵蚜危害时间增加表达量增加,在“新红星”中苹果绵蚜危害12 h时表达量最高,抗性最显著;MdOPR14基因在3个品种中抗性反应一致,随着苹果绵蚜危害时间增加表达量下降;MdOPR12,MdOPR13基因在“小国光”中随苹果绵蚜危害时间增加表达量下降,在“新红星”和“红富士”中随苹果绵蚜危害时间增加表达量先上升后下降,苹果绵蚜危害12 h时表达量最高;MdOPR5在“红富士”中随苹果绵蚜危害时间增加表达量下降. 总体而言,苹果树被苹果绵蚜危害后,MdOPR1和MdOPR4表达量显著增加,表明MdOPR基因可能通过调控JA的合成以响应苹果绵蚜对其生物胁迫.
 DownLoad:
DownLoad: