


 下载:
下载:
-
修枝即人为去掉林木活枝冠层的一种森林抚育管理措施,已在林木培育中广泛使用.国外已开展修枝对巨桉Eucalyptus grandis、大花序桉E. cloeziana、亮果桉E. nitens、细叶桉E. tereticornis和蓝桉E. globulus等树种的相关研究[1-5],国内也对史密斯桉E. smithii[6]、托里桉E. torelliana[7]和尾巨桉E.urophylla×E.grandis[8]进行了修枝试验.研究结果表明,对林木进行适当人工修枝,既能减少树体节疤,增加树干通直度和圆满度,又能促进林木生长、提高产量,进而培育出高干、优质无节材,同时还能抑制部分病虫害的发生[9-11].
大花序桉,美称澳洲大花梨,为桃金娘科桉树属昆士兰桉亚属Idiogenes树种,自然分布于澳大利亚昆士兰州[12],树高通常为35~40 m,最大高度40~55 m,直径1~2 m[13].木材黄褐色,纹理通直,结构均匀,沉重,坚固,硬度高,耐久.我国自1972年以来在广西、广东、福建等地广泛引种栽培,效果较好[14].大花序桉生长速度快,木材材质优,是很好的锯材树种,亦是培育中大径锯材的优良树种.本文拟对大花序桉幼林进行修枝试验,探索修枝对其生长量的影响,为培育优质大花序桉中大径锯材奠定基础.
全文HTML
-
试验林位于广西兴业六万林场,属亚热带季风气候,温暖湿润,光热充足,无霜期长,雨量充沛,平均气温为22.3 ℃,年极端最高气温为36.6 ℃,年极端最低气温为4.6 ℃.试验采用人工挖穴整地方式,株行距2 m×3 m,施用有效总养分25%的桉树专用基肥0.5 kg/穴,2013年7月完成造林,2014年5月施用有效总养分25%的桉树专用追肥0.5 kg/株,同时配施硼砂40 g/株. 2014年7月发生的超强台风“威马逊”给试验林造成严重损失,林木几乎全部倒伏,根系严重受损,经扶正后生长逐渐恢复.
-
试验设置4种修枝强度处理,修枝强度以修去活枝冠层长度占整个活枝冠层长度的比例表示,即分别为未修枝(S1);轻度修枝(修去活枝冠层的1/3下层枝条,S2);中度修枝(修去活枝冠层的1/2下层枝条,S3);重度修枝(修去活枝冠层的2/3下层枝条,S4).每个修枝强度对应一个标准地,每个标准地设置成10行×10株的试验区,按等高线将每个标准地自上而下设置为5次重复.于2015年3月30-31日完成人工修枝工作.
-
于2015年3月19日测定试验林的胸径和树高,2015年3月30-31日完成试验林的修枝工作,修枝完成后继续跟踪调查试验林的生长情况,分别于2015年7月11日和2016年3月29日对胸径和树高进行了测定.胸径采用围尺进行测定,数值单位为cm,精确到0.1 cm;树高采用瑞典生产的雷达树高测定仪进行测定,数值单位为m,精确到0.1 m.单株材积利用上述胸径和树高测定值进行计算,数值单位为m3,精确到0.000 1 m3.单株材积计算公式参见文献[15].本文分析了2015年3月-2016年3月胸径、树高和单株材积的定期生长情况,比较了不同修枝强度对生长的影响差异.
-
于2016年6月3日在4种修枝处理标准地中,分别选择生长较好的林木4株,使用5 mm口径生长锥在树干胸高处的南北向钻取一条贯通髓心的木芯,用胶管封装,避免水分蒸发.将木芯带回实验室,在实验台上把木芯从髓心处分为2段,其中一段由心材向边材方向较均匀地分为3小段,分别标记为L1,L2,L3.使用数显游标卡尺测定该3小段木芯的长度,利用圆柱体公式计算其体积,随后将木芯置入干燥箱中,将干燥温度设置为80 ℃,连续烘12 h,木芯的质量不再变化.将木芯放入装有干燥剂的干燥皿中,冷却后称质量,数值单位为g,精确到0.001 g.
木芯体积计算公式为
木材基本密度计算公式为
式中:V为木芯体积(cm3);d为木芯圆柱体半径(mm),本文d为2.5 mm;L为每段木芯长度(mm);D为基本密度(g/cm3);m为每段木芯绝干质量(g).
-
本文所有数据采用Excel 2007进行初步整理,并用Spss16.0进行数据分析.
1.1. 试验林概况
1.2. 试验设计
1.3. 试验方法
1.3.1. 生长测定
1.3.2. 木材基本密度测定
1.4. 数据分析
-
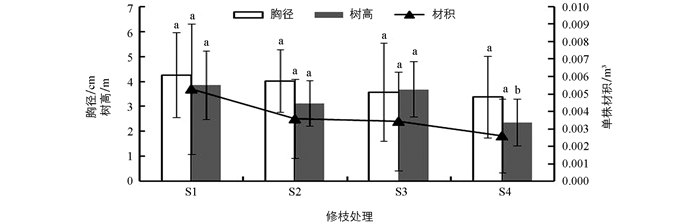
由表 1可知,不同修枝强度处理间的胸径和单株材积差异不具有统计学意义,但树高差异极具有统计学意义,说明修枝对大花序桉幼林的胸径总体变化影响不大,而对树高有较大影响,但二者共同作用下单株材积对修枝响应的差异不具有统计学意义.
由图 1可知,大花序桉幼林的胸径增长随着修枝强度的增加而逐渐下降,胸径增长最大的未修枝S1比重度修枝S4高20.73%;树高增长除了中度修枝S3比S2和S4大外,随着修枝强度的增加,其总体也呈现出下降趋势,而中度修枝S3的树高增长稍小于S1;单株材积随着修枝强度的增加而逐渐降低,其中轻度修枝S2和中度修枝S3相差不大,而增长最大的未修枝S1比重度修枝S4高43%.
-
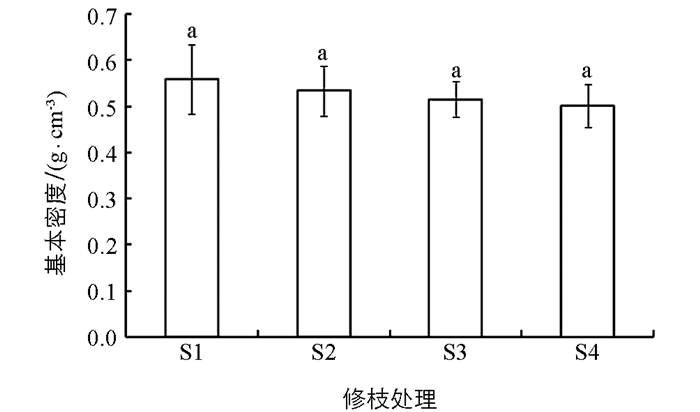
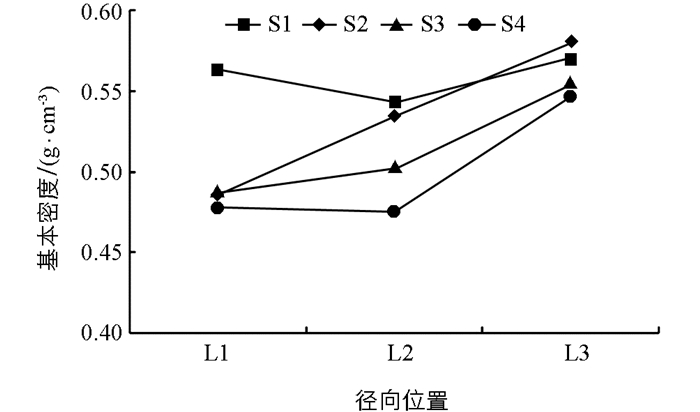
由表 2可知,不同修枝强度处理间的木材基本密度差异不具有统计学意义,说明修枝对大花序桉幼林木材基本密度的影响不大,但修枝引起基本密度的规律变化,随着修枝强度的增加,木材基本密度表现出递减趋势(图 2).不同径向位置基本密度的差异具有统计学意义,说明林木在生长发育过程中木材基本密度在径向上有差异性表现(表 2).不同修枝强度处理间径向位置的基本密度变化趋势如图 3,在未修枝林木髓心位置的基本密度大于修枝林木,同时修枝后林木髓心位置的基本密度基本一致.从径向位置中部L2的基本密度来看,修枝后林木的基本密度有所增加,而未修枝林木在下降.外部L3木材的基本密度都较大于中部L2和髓心位置L1,由此可见修枝明显降低了心材基本密度,对边材基本密度影响不大.与未修枝相比,修枝总体表现为降低木材基本密度的趋势.
-
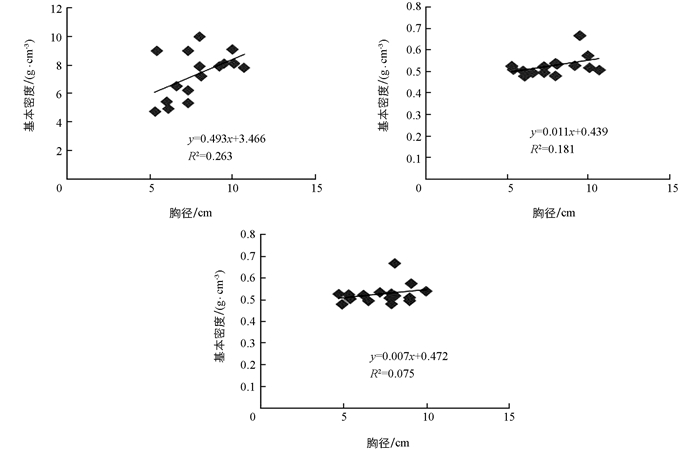
由图 4可知,树高与胸径成线性关系,但决定系数R2仅有0.263,说明在建立的线性方程中,胸径生长促进树高生长的贡献率也只有26.3%.胸径、树高与基本密度的线性关系较弱,决定系数R2分别仅有0.181和0.075.因此,胸径与基本密度、树高与基本密度的相关性不具有统计学意义.
2.1. 修枝对生长的影响
2.2. 修枝对木材基本密度的影响
2.3. 生长与木材密度的相关关系
-
对速生树种而言,修枝的主要目的在于不降低或提高林木生长量的同时,改善树体外观质量,提高木材材质,增加木材附加值.本文修枝对大花序桉胸径生长的影响虽然表现为差异不具有统计学意义,但却呈现出胸径增长随修枝强度增加而降低的趋势,这与任世奇等[8]对尾巨桉修枝后胸径定期生长量影响的研究结果一致,说明大花序桉修去的枝条仍在进行正向同化作用,生成有机物,促进林木生长.当修掉下层活枝后,由于有机物质的就近分配原则,上层活枝生产的有机物不能送达到树体下部.因此,修枝对胸径增长的影响差异不具有统计学意义.树高增长除中度修枝S3稍小于未修枝S1外,轻度修枝S2和重度修枝S4较未修枝S1表现为递减趋势,与吴际友等[16]对火炬松修枝后树高变化效应一致,说明轻度修枝S1修掉了少量下层活枝后,轻微降低了整树有机物的同化量,使轻度修枝S1的树高增长稍小于未修枝S1,而重度修枝S4因修掉过多的枝条而严重降低了整树有机物的同化量,使树高增长降低最多.对于中度修枝S3稍小于未修枝S1,大于轻度修枝S2和重度修枝S4,可能正如任世奇等[15]和牛正田等[17]所述,修掉过多的枝条,对单株的伤害刚好超出了特定的临界点而激发了增强生长维持功能,提高了单叶的光合能力,进而维持了有机物的同化量.单株材积由胸径和树高共同决定,本文将单株材积作为衡量修枝对大花序桉幼林生长较适宜的强度为中度修枝S3,即是修掉活枝冠层的1/2,这与任世奇等[8]对尾巨桉的研究结果一致,说明大花序桉和尾巨桉同属桉树属树种生长对修枝有一致的响应.
本文修枝对木材基本密度影响的差异不具有统计学意义,这与Tong等[18]对北美短叶松修枝前后木材密度研究结果一致,但随着修枝强度增加木材密度呈现出递减趋势.不同径向位置的木材密度从髓心处向外部逐渐增大,其中重度修枝S4的斜率最大,增大的幅度最大,说明修枝对后期生长的木质部密度有促进作用.此外,胸径与树高的线性相关决定系数仅有0.263,说明二者的相关性不显著;胸径与基本密度、树高与基本密度的线性相关决定系数均非常小,说明他们的相关性较差,这与任世奇等等[19]的研究有所不同,其原因可能是本研究试验林在种植1年后遭受超强台风,导致树体受损严重,对本研究结果造成了一定影响.因此,本研究将持续跟踪观测,以进一步探索修枝对大花序桉生长的长期作用.