


 下载:
下载:



-
魔芋是天南星科(Araceae)魔芋属(Amorphophallus)多年生草本植物,是迄今发现的极少能大量合成葡甘聚糖(konjac glucomannan,KGM)的植物种群,主要分布在亚洲和非洲热带及亚热带的一些国家和地区,仅部分亚洲国家有栽培.大面积主栽种为花魔芋(A.konjac K. Koch)和白魔芋(A.albus P. Y. Liu et J. F. Chen),其中我国学者刘佩瑛、陈劲枫发现并命名的新种白魔芋为我国特有,品质最佳.魔芋是我国传统栽培的重要经济作物,具有极高的经济价值,其所含葡甘聚糖具有诸多优点,已成为食品、医药、化工、石油、纺织等工业中具有重要用途的原料与产品,出口欧美、日本等,带动了一大批相关产业的发展[1].
葡甘聚糖含量是魔芋品质的最重要衡量指标. 1975年,Muoat通过实验认定GDP-甘露糖是魔芋葡甘聚糖生物合成的甘露糖供体,而GDP-甘露糖的合成由GDP-甘露糖焦磷酸化酶催化形成,因此GDP-甘露糖焦磷酸化酶对于葡甘聚糖的合成起着重要调控作用.何春梅等的研究表明[2],在铁皮石斛中,GDP-甘露糖焦磷酸化酶参与葡甘聚糖的合成、抗坏血酸合成以及糖基化作用等过程,对植物的正常生长以及抗逆性起着重要作用[3].近年来,已有学者在其他一些植物例如拟南芥中分离纯化出这种酶,得到了GDP-甘露糖焦磷酸化酶基因的全长序列,并对其功能进行了研究[4].但在魔芋当中,对于GDP-甘露糖焦磷酸化酶及其基因的研究非常少,仅有Markus Pauly[5]通过转录组454高通量测序方法,得到该基因的全长序列,并已在GenBank登陆(登陆号JF727269).其基因全长801 bp(包含部分UTR区域),编码氨基酸长度是146个(登陆号AEH27534.1),对比其他物种GDP-甘露糖焦磷酸化酶氨基酸长度为360多个氨基酸,数目和序列相差都很大.且文中未显示该序列的PCR扩增鉴定,更未有进行功能验证,故Markus Pauly得到的GMP基因序列可能存在一定的问题.本研究将重新克隆该基因及其启动子序列,将得到的基因序列与其他物种进行对比,并对启动子序列进行功能分析预测,以期初步验证我们的猜想,也为之后进行该基因的功能验证打下基础.
全文HTML
-
花魔芋、白魔芋叶片、球茎等.
-
大肠杆菌Trans1-T1 Phage Resistant Chemically Competent Cell和克隆中间载体pEASY-Blunt Simple Cloning Kit购自全式金生物;克隆中间载体pClone007 Simple Vector Kit购自北京擎科新业生物技术有限公司;
-
Quick RNA Isolation Kit试剂盒(北京华越洋生物技术有限公司);TaKaRa反转录试剂盒(宝日医生物技术有限公司);克隆所需酶Phanta Max Super-Fidelity DNA Polymerase和2×Taq Master Mix(诺唯赞生物);Trans2K Plus Ⅱ DNA Marker(全式金生物);高效植物基因组DNA提取试剂盒Hi-DNAsecure Plant kit、通用型DNA纯化回收试剂盒Universal DNA Purification Kit、质粒小提试剂盒TIANprep Mini Plasmid Kit(天根生化科技有限公司);Nucleic Acid Stain核酸染料(北京鼎国昌盛生物技术有限责任公司);PCR引物合成和DNA测序由北京擎科新业生物技术有限公司完成,X-Gal,LB,琼脂粉等为常规试剂.
-
利用Quick RNA Isolation试剂盒提取花魔芋球茎总RNA,利用TaKaRa反转录试剂盒将RNA反转录成单链cDNA,并通过魔芋仅有的5个转录组高通量测序数据库,与拟南芥中GMP氨基酸序列,进行Blast比对、拼接,得到魔芋GMP基因的推定全长,尽量包括UTR区域,再据此序列设计基因特异性引物GMP-F/GMP-R(表 1).
使用高保真酶进行GMP基因的克隆,PCR反应程序为:95 ℃预变性3 min,95 ℃变性30 s,60 ℃退火15 s,72 ℃延伸50 s,32个循环,总延伸5 min. PCR产物经1%琼脂糖凝胶电泳检测后回收目标条带,将纯化后的产物与pEASY-Blunt Simple Cloning载体连接,再经过42 ℃热激转入大肠杆菌Trans1-T1 Phage Resistant Chemically Competent Cell中,菌落PCR检测后将阳性克隆送北京擎科新业生物技术有限公司测序.
-
以魔芋幼叶为材料,利用试剂盒提取花魔芋的总DNA.根据已克隆得到的GMP基因序列测序结果,利用FPNI-PCR法克隆GMP基因的启动子,设计3个嵌套式的特异性引物80063,80064,80065(表 1),3轮反应程序具体参照WANG等[6]的方法,在第3轮中发现3条特异条带,经切胶回收、热激连接pClone007 Simple Vector,阳性菌落检测,送样测序.
根据第1轮测序结果设计引物,分别在距离ATG上游2 116 bp和207 bp处设计扩增全长启动子上下游引物,为80073和80065(表 1).以花魔芋的总DNA为模板,使用Phanta Max Super-Fidelity DNA Polymerase进行扩增,PCR反应程序为:95 ℃预变性3 min,95 ℃变性15 s,57 ℃退火15 s,72 ℃延伸1 min,30个循环,总延伸5 min.经切胶回收、热激连接pEASY-Blunt Simple Cloning载体(全式金生物),阳性菌斑检测,送样测序.
利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线数据库分析所克隆得到的GMP基因的启动子调控原件,据此,对GMP基因的启动子进行功能分析预测,利用TSSP(http://www.softberry.com/cgi-bin/programs/promoter/tssp.pl),分析预测GMP基因转录起始位点.
1.1. 材料
1.2. 菌株和载体
1.3. 试剂
1.4. GDP甘露糖焦磷酸化酶基因的克隆
1.5. GDP甘露糖焦磷酸化酶基因启动子的克隆和其生物信息学分析
-
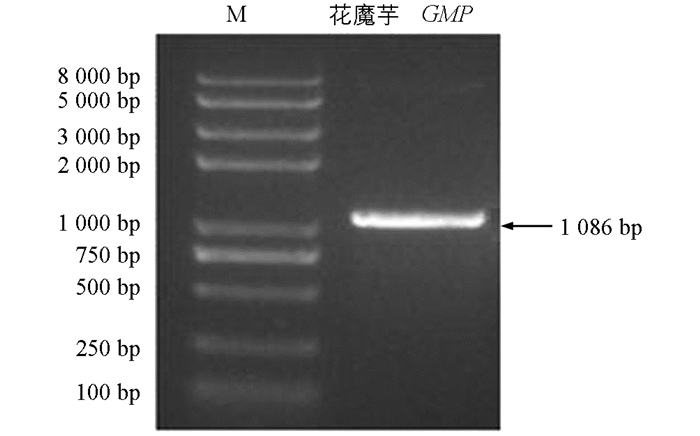
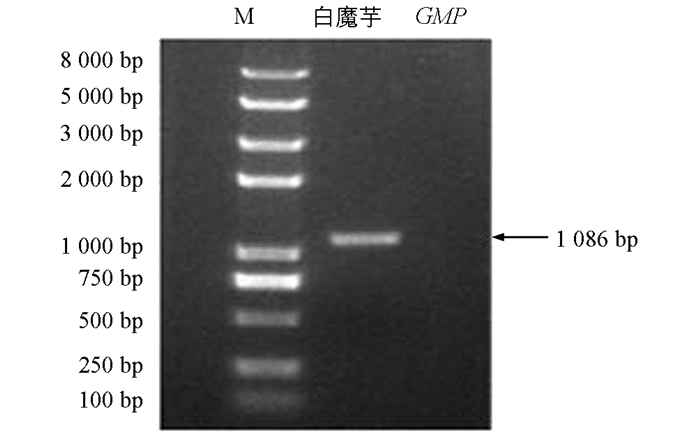
通过仅有的魔芋5个转录组高通量测序数据库拼接得到魔芋GMP的推定序列,设计基因特异性引物扩增GMP基因,测序结果发现,得到了花魔芋和白魔芋中部分cDNA序列:CTCTCCTCGTTGGGATCTGCTGTCGGCGCATCCAATCCGGGGCATCTCCAGTAGGGCACGAGAAGGGAGGGTTAGCATCCCGCCCACCATGAAGGCACTCATCCTGGTCGGAGGCTTTGGCACTCGGCTACGGCCGCTGACACTCAGCGTCCCCAAGCCGCTTGTCGATTTCGCCAACAAGCCCATGATCGTACACCAGATCGAAGCTCTCAAAGCAATTGGAGTCACAGAAGTGGTCTTGGCCATCAACTACCAACCAGAGGTAATGCTCAACTTCTTGAAGGAGTACGAAGCCAAGCTGGGGCTCAAGATCACCTGCTCTCAGGAGACTGAACCACTCGGGACGGCCGGTCCCCTGGCCCTGGCCAGGGACAAGCTGATAGATGACTCCGGCGAGCCCTTCTTTGTCCTCAACAGCGATGTCATCAGCGAGTACCCCCTCGCTGAGATGATCCAGTTCCACAAGTCTCATGGGGGGGAAGCATCCATCATGGTGACCAAGGTGGATGAGCCTTCAAAGTATGGGGTGGTTGTTACGGATGAGGAGACCGGGAAAGTAGAGAAATTTGTGGAGAAGCCAAAGATCTTTGTGGGCAACAAGATCAACGCCGGCATCTACCTGTTGAACCCCTCTGTTCTGGACCGGATCCAGCTCCGTCCCACCTCCATCGAGAAGGAGGTCTTCCCCAAGATCGCCGCGGAGAAGCAGCTCTATGCCATGGTCCTCCCCGGGTTCTGGATGGACATCGGCCAGCCGAGGGACTACATCACCGGCCTGCGCCTCTACCTCGACGCCCTCCGGAAGCGGTCTGCCGCCAGGCTCGCCGTCGGCCCCCACGTCGTCGGCAACGTTTTGGTGCACGAGACGGCGGCCATCGGGGAGGGCTGTCTGATCGGCCCCGACGTGGCCATCGGCCCGGGCTGCGTCGTCGAGCCCGGCGTCCGGCTGTCGAGGTGCACGGTGATGCGCGGGGCCCGCATCAAGAAGCACGCCTGCGTCTCCAGCAGCATCATCGGGTGGCACTCCACTGTTGGGCAGTGGGCGCGGGTGGAGAACATGACCATCCTCGGGGAGGACGTGCATGTGGGCGATGAGATTTACAGCAATGGCGGGGTGGTGCTCCCCCACAAGGAGATCAAGTCCAGCATATTGAAGCCTGAGATCGTCATGTGATGTGAGTGGCCCAAGTCCATGTTGTCCATGTCACTGTTCCAAGCTGCGTCCTCTCTCTC,长度为1 233 bp,推定开放阅读框(Open Reading Frame,ORF)为1 086 bp,编码361个氨基酸残基,且两种魔芋GMP的氨基酸序列一致(图 1和图 2).
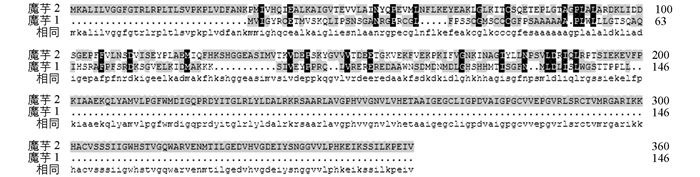
利用DNAMAN,将Markus Pauly登陆的花魔芋GMP蛋白的氨基酸序列与拟南芥、木薯、大桉中的GMP蛋白的氨基酸序列进行比对(图 3),发现氨基酸的长度与序列相差很大,同源性很低.而将本研究得到的GMP蛋白的氨基酸序列与拟南芥、木薯、大桉中的GMP蛋白的氨基酸序列比对(图 4),GMP蛋白与拟南芥(NP_181507.1)中GMP蛋白(NP_181507.1)具有88%的同源性,与木薯中GMP蛋白(XP_021626075.1)具有93%的同源性,与大桉中GMP蛋白(XP_010067451.1)具有92%的同源性.本研究得到的魔芋的GMP蛋白的氨基酸序列与Markus Pauly所登陆的GMP蛋白的氨基酸序列差距很大(图 5).因其在论文中并没有提及对GMP基因进行功能验证,我们猜想Markus Pauly得到的GMP基因全长序列存在一定的问题,并经过以上分析,认为我们更有可能得到的是魔芋中GMP基因的正确全长序列.
在魔芋和其他物种基因组中对GMP的相似基因进行分析,利用MEGA7.0基于最大似然法的分子系统发育分析构建含有13个GMP氨基酸序列的进化树(图 6),用于建树的酶如下:拟南芥AtGMP(NP_181507.1)、木薯MeGMP(XP_021626075.1)、大桉EgGMP(XP_010067451.1)、橡胶HbGMP(XP_021653979.1)、芝麻SiGMP(XP_011076786.1)、阔叶猕猴桃AlGMP(ACN38266.1)、哈密瓜CmGMP(XP_008441398.1)、莲NnGMP(XP_010256792.1)、南瓜CmGMP(XP_022954947.1)、榴莲DzGMP(XP_022765102.1)、醉蝶花ThGMP(XP_010554624.1)、大豆GmGMP(ACW84415.1).结果表明,我们所得到的GMP与其他植物的GDP-甘露糖焦磷酸化酶的系统发育有一定的相关性,这进一步显示了本研究得到的魔芋GMP基因的正确性.
-
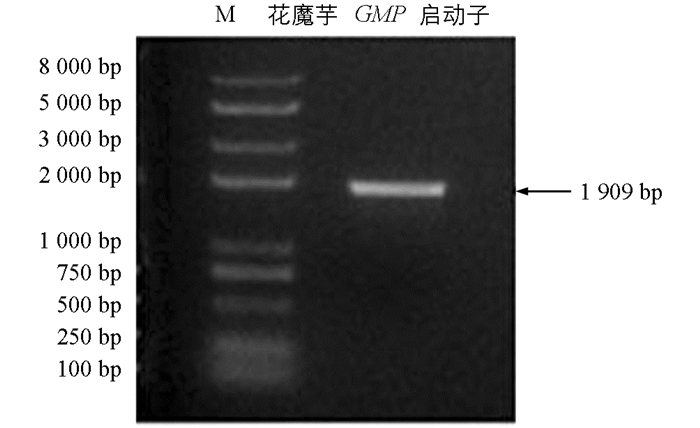
利用染色体步移技术FPNI-PCR方法对GMP启动子序列进行两次克隆.第1次使用普通Taq酶扩增,PCR扩增产物存在3种条带的大小,经测序验证分别为3种长度的该基因的起始密码子ATG上游序列,即启动子序列,其中最长的启动子序列接近2 000 bp,因此第2次扩增使用高保真酶,以第1次扩增得到最长启动子序列为模板,在距离ATG上游2 116 bp和207 bp处设计上下游引物,最终在花魔芋总DNA中克隆得到1条长1 909 bp的片段(图 7),加上后面还有ATG上游的207 bp,拼接得到了ATG上游2 116 bp的启动子序列:CATAGGGAGGGCGATAGAGGGGGAGGGGGCATGCACCAACGGTGGGGGGAGAGGGAGCCAGCGGTGAGAGAGAGAGGGCGAGGAGGGTCTACTACGAGGTGGAGGAGAGAGAGAGGGGGAATGCGCTCTCTCTCTCTCTTAGTAAAGGTATTTTTAAAATATTCTACAAAATATTTAACTCCTATCGACGGTAGTAAATTATAATAGCATAACCACGACATTTAAGACTACACAGAGGATATTTTTGATAATATATTCAAATGAAATATATATTCGATATAACCGATACTCGGAGAGATATTTATATAAAAATTTAAAAGATGAGAATTTTTTGAGGCGCAGTATGCCACGGTGTAACCGCCAATACACGCCTTTACCGGGTGCGGACGCGAGCCGATTCCTTAAGCCGGGGAAAGCGCCGGTTTGACACCCCACGTCGAAGAAAGTTGGGGGGGGGGAGGGGGGAAAAATAAATATTAAAAGTAAGGAAAACGTTTCCGAAATGACTCCACCCTGGTTGGCATGTAGCGGAAGAGAGATGAGAGAGAAAATATAGAGGGAGAGAAAGATGAGGAGTGAGAGAGACACATGAGATAGAGAGAAGAGAGAATAAGAGATGACGACGATGACGATGGGGGGGGAGAGAGAGAGAGAGGTAGGGAGAGAAGAGAACAGTGGGGTGGTGTCGCTGTGGAGATCGGGGGAAGGCTATAAAACCCGTGGCAGCGGGGCTGAGTTCGCAGCACAGACCGTTGGCGGGACTCGGGAGGACGTGCTCGTCATTTCCACGTCTCTCTGCTCCTCCTCACACACCTCTCTCTCTCTCTCTCTCTCTCTCTAACACACCCGCTCCCTTCCTCTGTCTTCTTCTCCCGAGAGGGAGAGCGCGAGGAGAAACAAGCCCGCGGGGTGGGCGGGATCCAAGTCGCGGGCTTTTGCCTCCGCAGTCCGCCGGGACTCCATCTCGGTAAACCACCGCATCCCGCCTGCACGCTCTTCTCTCCCGGTTTCCGTGCCTTTTTCTACTGCATTCGCCCGTTTTCCGCCGGATCTGGAGGGGGAGACCCGGCAACGTAGCCGGTTTCTTCCCTCGCCTTTGTTGGCTTCTGGGATCTCGAGGGGGAGCCCGGCAATGGCCAAGGGAAGAAGCTTCTTTGCGCGTCTCCTGCGCTGGATCCCAGAAGCTATGCTTTCTTTCCCAATCCCATGCCATCGCCGTCACCGAATTGCTCTGCTGACACCTGGTTTGGGCGGCGGCGTTCTGTTTTACCGGTCCCTTTTTCCACTCCCCCGGATTTGGGTGCCTTTGGAATCTCTGTTTCCGGGCGTTTTATCCCCATCCCGATGCTTATGAGGATCGTCCATTGCTTGTAATGAGGTAGATCTGATAAATTCATAACCTTTTCCCTTTGCTAGCGGCCAAACGCCAAGAACAGGACATATCAGCAACTTTTTTTCTGTATCCCCTCTTCCCCGGGCTTCCCATTTCACGCATGGGGCTCTCTTCTTCCAGCTCTTTATCCGAATTGGTTAGAATAGAACCGGACAGACGGACTAGGTGAAGAAAGGGGAACAGAGGACTCCAACTGGCTCCTTTGGCTTCTGCAATAGAAGACGATTCCAGCTTGCAGTGTAGCCGGAAAGATTTTGTTCCCGAGCTCCAGCTAGAACGTATGAAGACACATTTGTCCTGTCACTGCCAAGAGGGTTTTAGGCGTCTGCGGACACTTGTTGGGGCATTTCAGTCAAGCAGCTGAAACCCCTCTCTTTGTAAAATCCCGCCCTCGTCCTCCCAACGGTTTCCCAGCTCCTCTCCTCGTTGGGATCTGCTGTCGGCGCATCCAATCCGGGGCATCTCCAGTAGGGCACGTAAGGTTCATGGCCTTTGTGCTTTTGTGTGGCAGAAGCATCCCGTCATAGCTTGATAGCCTTCAGCTTACACATGGCAGGCTTCTCCATGGAGGAGCTTCTCAGCTTCCCACCACTCCGTAGCAGGAATGACATGAGATTCTGCACGCATCTATATATGGCTTTGCAGTTCTGGGTGGCGCCGTGAGATTAACCTGTGTTCTGTTTCTGATCTGCAGGAGAAGGGAGGGTTAGCATCCCGCCCACC.
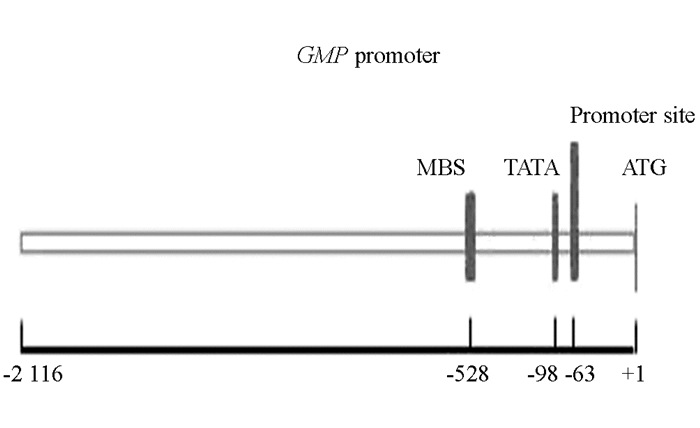
利用PlantCARE在线分析软件对GMP基因启动子序列进行功能分析预测(表 2).利用TSSP在线分析软件预测GMP启动子的转录起始位点(图 8).该启动子除了典型的TATA-box和CAAT-box核心启动子原件外,还包含了多种光应答、光响应元件、激素响应元件.值得注意的是,除了光应答、激素响应元件之外还存在一些其他的环境响应元件,例如厌氧诱导所必需的顺式作用调节元件ARE,抗旱和高温响应元件CCAAT-box、MBS等.另外,还发现了参与细胞周期调控的顺式作用元件(MSA-like),胚乳表达所需的顺式作用调控元件(Skn-1_motif),参与水杨酸反应的顺式作用元件(TCA-element)等.可以推测GMP基因的启动子可能受水杨酸(SA)、茉莉酸甲酯(MeJA)、脱落酸(ABA)诱导调控,还可能参与抗逆反应的调控,需要进一步验证.
2.1. 魔芋GMP基因克隆
2.2. 花魔芋GMP启动子克隆和顺式原件预测
-
由于与魔芋的近缘物种,即使是一个科的研究都很少,魔芋尚未进行全基因组测序,目前魔芋分子生物学特别是功能基因组的研究相当滞后,已发现的基因组资源极为有限,在GenBank上登陆的魔芋属(Amorphophallus)的核酸序列、ESTs和克隆的基因序列都很少,而关于葡甘聚糖合成调控分子机制的研究更是有限,这都极大地阻碍了魔芋品质改良的进程. GDP-甘露糖焦磷酸化酶(GMP)直接控制GDP-甘露糖的合成,为魔芋葡甘露聚糖的合成最后一步提供两个底物之一,故是魔芋葡甘露聚糖生物合成路径中的关键酶.目前已有学者从一些植物中克隆得到了GMP基因并对其进行了功能验证,如拟南芥、铁皮石斛.在魔芋中,对于GDP-甘露糖焦磷酸化酶及其基因的研究非常少,仅有Markus Pauly[5]通过转录组454高通量测序方法,得到注释为花魔芋的GDP-甘露糖焦磷酸化酶基因的全长序列,并已在GeneBank登陆(登陆号JF727269).登陆的基因全长801 bp(包含部分UTR区域),编码氨基酸长度是146个,对比其他物种GDP-甘露糖焦磷酸化酶氨基酸长度为360多个氨基酸,数目和序列相差都很大.但其研究结果未显示该序列的PCR扩增鉴定以及功能验证,我们推测所登陆的该基因序列很可能存在问题.因此对魔芋GDP-甘露糖焦磷酸化酶基因进行克隆和功能研究是非常有必要的.本研究利用魔芋的转录组高通量测序数据库,进行tBlastn和拼接,再据此设计基因特异性引物,扩增得到推定的GMP基因,包含部分UTR区的序列,其编码氨基酸序列与其他物种的具有很高的相似性,初步说明我们得到的应该是该基因,关于其功能验证正在进行中.
现阶段克隆启动子比较常用的方法有PCR法、TAIL-PCR[7]、接头PCR[8]等方法. PCR法较为简便,但只限于已知序列的克隆;TAIL-PCR近年来应用比较广泛,但过程较为繁琐,总之这些启动子克隆方法各有利弊.由于魔芋的整个基因组没有进行完全的测序,因此花魔芋GMP基因的启动子序列是未知的,不能直接用常规的PCR的方法进行克隆,本研究采用了FPNI-PCR的方法[9]克隆得到了GMP基因ATG上游长度为2 116 bp的启动子序列,并在拼接得到的启动子序列的基础上设计引物,运用常规的PCR方法,使用高保真酶,在花魔芋总DNA中再次克隆得到GMP基因的启动子序列.运用PlantCARE等在线软件对序列进行预测分析[10],发现该启动子含有多种生物学顺式作用元件,推测GMP基因的转录水平可能受到多种因素的调控.除了典型的TATA-box和CAAT-box核心启动子原件外,还具有一些与光响应、激素相关的调控元件,推测GMP的转录水平与激素的调节有关.在拟南芥中,已研究得到GDP-甘露糖焦磷酸化酶参与抗坏血酸的合成;在铁皮石斛中,也有研究表示该酶与抗盐胁迫密切相关.因此,我们推测在GMP基因的启动子中可能含有一些与抗胁迫相关的调控元件.通过对得到的启动子序列的分析发现,除以上顺式作用元件外,该序列中还包含MYBHv1结合位点元件,厌氧诱导所必需的顺式作用调节元件等非生物胁迫响应元件,暗示在魔芋中GMP基因可能参与对非生物胁迫的耐受能力.
已有研究表明,GDP-甘露糖焦磷酸化酶参与植物多糖合成、抗坏血酸合成以及糖基化作用等过程[11],对植物的正常生长以及抗逆性起着重要作用.鉴于Markus Pauly虽拼接得到花魔芋GMP基因的序列,但是并没有对其进行功能验证,而本研究得到的魔芋GMP基因的全长序列具有较强的保守性.并且本研究还克隆得到了该基因的启动子序列,该序列具有可能提高植物对非生物胁迫的耐受能力的调控元件,因此GMP基因极有可能与抗逆性有关,这更加验证了我们的猜测——Markus Pauly在GeneBank中登陆的GMP基因的全长序列存在一定的问题.为了证实我们的猜测,还需要进行GMP基因及其启动子的功能验证[12]等方面的研究.