


 下载:
下载:
-
北美红枫(Acer rubrum L.)为槭树科(Aceraceae)槭树属(Acer L.)大型落叶乔木.原产北美洲东海岸,因其树形优美,叶色多彩,近年来被广泛引种于国内[1].北美红枫树高可达30 m,冠幅可达12 m,寿命可达100年.其适应能力强,生长速度快,易成形成林,非常适合作景观树种,为城市增添美感[2].北美红枫变种达40多种,在抗寒性、生长速率、秋季着色和叶片形态等方面表现出高度差异[3].目前,国内外学者对北美红枫的研究主要针对单一品种对环境条件的响应[4]、扦插育苗[5]、组培快繁[6]和栽培管理[7]等方面.李力等基于Lab模型对其呈色生理机制进行过探讨[8],指出花色素苷、光合色素和可溶性糖是影响叶片a值的重要因素,喷施蔗糖溶液不会显著影响叶片转色[9].研究发现,在秋季变色期,被引进到重庆地区的秋火焰(Acer rubrum ‘Autumn Blaze’)和酒红(Acer rubrum ‘Brandy Wine’)的叶片多数在转红前便已脱落,而十月光辉(Acer rubrum ‘October Glory’)的叶片转红早,效果好,持续时间长.为明确北美红枫的呈色机制,本试验以秋火焰等3种北美红枫为材料,测定和比较呈色期叶片的色素质量比、可溶性糖质量比、可溶性蛋白质量比、相关酶活性和叶片pH值等生理指标,研究北美红枫呈色的生理机制,以期为人工调控北美红枫秋季叶色提供理论依据.
全文HTML
-
试验地位于重庆市北碚区西南大学后山竹园实验基地(29°49′2.4″N,106°24′56.0″E).试验地土壤基本理化性质测定结果为:紫色土,pH值为5.9,全氮1.39 g/kg,有效磷14.93 mg/kg,速效钾20.31 mg/kg,有机质22.46 g/kg,容质量1.334 g/cm.北碚地区气候类型为亚热带季风气候,秋季降雨日较多.试验期间当地最高气温34 ℃,最低气温12 ℃.
-
试验材料为从美国引进的3种北美红枫:秋火焰、酒红和十月光辉.每个品种选取生长良好、长势相近的植株4棵(平均地径6 cm、平均株高4.0 m、树龄5年).从2016年9月30日至2016年11月5日每隔9 d采样一次.采样时选取各样本树东、南、西、北4个方位,各方位各选取3片等高、健康、无损伤的叶片,立即带回实验室进行相关测定.每次采样前一天,各样本树随机选择3根枝条调查红叶、黄叶、绿叶比例,并用其描述3种北美红枫变色情况,变色情况记录见表 1.
-
CIE Lab颜色空间是由国际照明委员会(International Commission Illumination)于1976年制定的一种色彩模式,常作为均匀色彩空间的近似[10].它用3个基本坐标L*,a*,b*,本文省略*,简写为L,a,b,其中L代表明暗;b代表黄蓝;a代表红绿.通过L,a,b值可定量描述3种北美红枫秋季变色期叶色差异.叶色参数的测定参照李力的方法[8].将所采叶片固定,并用参数为ISO 500、快门320、光圈F4.5的Nicon D7000相机拍照,在Adobe Photoshop CS3拾色器中,采用Lab颜色模式,记录各个叶片的L值、a值及b值,然后求其平均值.
-
花色素苷质量比测定参照Lee等的方法[11].称取1.0 g新鲜叶片并剪碎,将其移至25 mL具塞试管,用移液管向具塞试管中加入1%盐酸乙醇10 mL,摇动试管至叶片碎屑全部浸没,避光置于32 ℃恒温水浴锅中4 h获得粗提取液,将粗提取液用离心机在5 000 r/s下离心10 min,其所获上清液即为花色素苷提取液.每个处理均设置3个重复.
-
叶绿素质量比和类胡萝卜素质量比的测定参照邱念伟等的方法[12-13];PAL活性参考刘金、Lister等的方法[14-15]测定;PPO活性参考Anderson的方法[16]测定;可溶性糖、可溶性蛋白质量比测定方法参照《植物生理生化实验原理与技术》[17],分别选用蒽酮比色法、考马斯亮蓝G-250染色法;叶片pH值的测定参考Lee等的方法[18].
-
试验数据方差分析及相关性分析选用SPSS 21.0软件,在两两比较差异是否具有统计学意义前,先对方差进行同质性检验,若可以认为齐性,则用LSD方法;若不能认为齐性,则用Tamhane’s T2(M)方法;对各因子进行相关性分析时,选用3种北美红枫平均值.选用Origin2017软件进行绘图.
1.1. 试验地概况
1.2. 试验材料
1.3. 试验方法
1.3.1. 叶色参数测定
1.3.2. 花色素苷质量比测定
1.3.3. 其他生理指标测定
1.4. 数据分析
-
由表 2知,3种北美红枫L值、b值均呈下降趋势,a值均呈上升趋势.整个秋季变色期,秋火焰、酒红、十月光辉的L值分别下降19.5,32.72,31.75;b值分别下降23.06,29.62,16.53;a值分别升高43.75,36.01,47.98. 11月5日,三者a值均达到峰值,此时十月光辉a值为秋火焰的1.67倍,是酒红的2.61倍.秋火焰11月5日a值仅比十月光辉10月18日a值大1.19,酒红11月5日a值甚至比十月光辉10月18日a值还要小5.84.十月光辉L值、a值始终大于秋火焰、酒红,且与二者的差异具有统计学意义.除11月5日外,秋火焰、酒红b值差异均无统计学意义. 10月18日十月光辉a值转正前,3种北美红枫b值的差异均不具有统计学意义,且十月光辉b值最小. 10月18日十月光辉a值转正后,其b值始终大于秋火焰、酒红,且与二者的差异具有统计学意义.
-
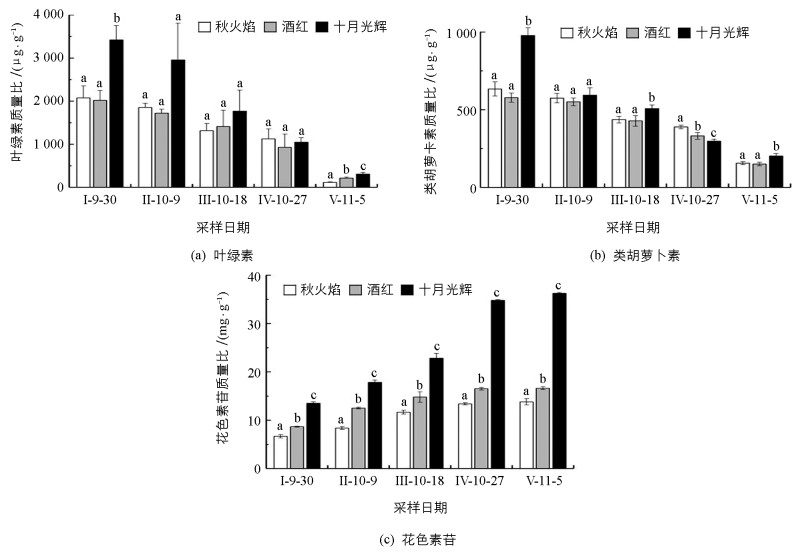
整个秋季变色期,3种北美红枫叶绿素、类胡萝卜素质量比均呈下降趋势,花色素苷质量比均呈上升趋势(图 1).十月光辉叶绿素质量比始终高于秋火焰、酒红;9月30日,十月光辉叶绿素质量比是秋火焰的1.65倍,是酒红的1.69倍;11月5日,十月光辉叶绿素质量比是秋火焰的2.68倍,是酒红的1.43倍.从9月30日到11月5日,秋火焰、酒红、十月光辉叶绿素质量比分别下降了1 959,1 805,3 114 μg/g(图 1(a)).
除10月27日外,十月光辉类胡萝卜素质量比始终显著高于秋火焰、酒红;9月30日,十月光辉类胡萝卜素质量比是秋火焰的1.54倍,是酒红的1.69倍;11月5日,十月光辉类胡萝卜素质量比是秋火焰的1.30倍,是酒红的1.34倍.从9月30日到11月5日,秋火焰、酒红、十月光辉类胡萝卜素质量比分别下降了477.35,426.60,775.63 μg/g(图 1(b)).
3种北美红枫花色素苷质量比均呈上升趋势,且三者差异始终具有统计学意义,花色素苷质量比从高到低顺序为:十月光辉、酒红、秋火焰. 9月30日,十月光辉花色素苷质量比是秋火焰的2.02倍,是酒红的1.56倍;11月5日,十月光辉花色素苷质量比是秋火焰的2.63倍,是酒红的2.18倍.从9月30日到11月5日,秋火焰、酒红、十月光辉花色素苷质量比分别增加了7.11,7.96,22.71 mg/g(图 1(c)).
10月18日后,3种北美红枫叶绿素、类胡萝卜素质量比差异相对较小,花色素苷质量比差异极大.
-
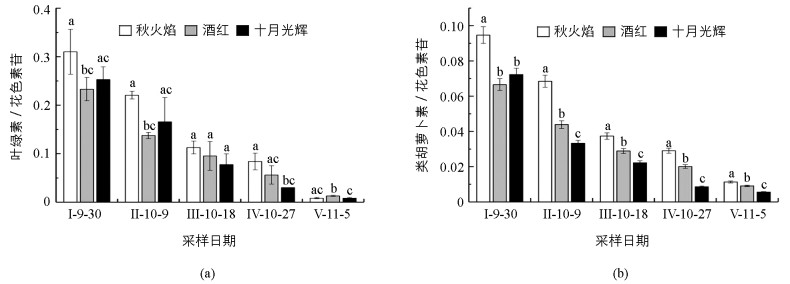
整个秋季变色期,3种北美红枫秋季变色期叶绿素与花色素苷质量之比(mChl/mAnt)、类胡萝卜素与花色素苷质量之比(mCar/mAnt)均呈下降趋势(图 2). 11月5日mChl/mAnt值与9月30日相比,秋火焰下降了37.36倍,酒红下降了17.93倍,十月光辉下降了29.76倍;10月18日后,十月光辉mChl/mAnt值始终小于秋火焰、酒红(图 2(a)). 11月5日mCar/mAnt值与9月30日相比,秋火焰下降了8.37倍,酒红下降了7.35倍,十月光辉下降了12.95倍;10月9日后,3种北美红枫mCar/mAnt值差异始终具有统计学意义,且十月光辉mCar/mAnt值始终小于秋火焰、酒红(图 2(b)).
-
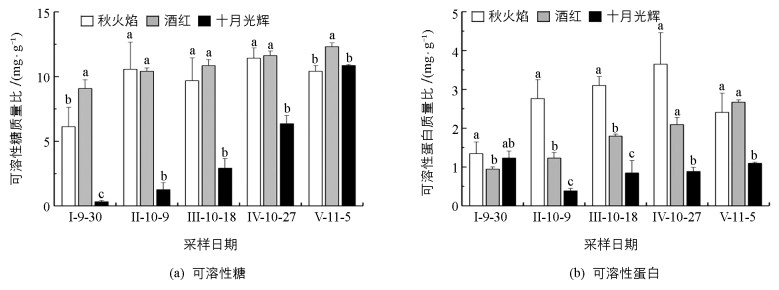
整个秋季变色期,3种北美红枫可溶性糖质量比(简写为SS)总体基本呈增长趋势. 11月5日采样结果与9月30日相比,秋火焰、酒红、十月光辉SS质量比分别上升了70%,36%,3 200%,十月光辉SS质量比极大提高.十月光辉5次采样的SS质量比比例约为1:4:9:19:33,接近为幂指数增长. 9月30日,秋火焰、酒红、十月光辉三者SS质量比接近为19:27:1,三者差异极具有统计学意义.而后至10月27日,秋火焰、酒红SS质量比仍显著高于十月光辉. 11月5日,十月光辉SS质量比达到峰值,虽略高于秋火焰,但仍显著低于酒红(图 3(a)).
整个秋季变色期,秋火焰可溶性蛋白质量比(简写为SP)呈先升后降趋势,酒红SP质量比基本呈直线上升,而十月光辉SP质量比则呈先降后升趋势. 10月9日,秋火焰、酒红、十月光辉SP质量比差异具有统计学意义,比例约为7:2:1. 10月18日,三者SP质量比均有所提高,且差异仍具有统计学意义,比例约为4:2:1. 9月30日至11月5日,3种北美红枫可溶性蛋白质量比按从高到低顺序大致为:秋火焰、酒红、十月光辉. 10月9日后,十月光辉可溶性蛋白质量比始终显著低于秋火焰、酒红(图 3(b)).
-
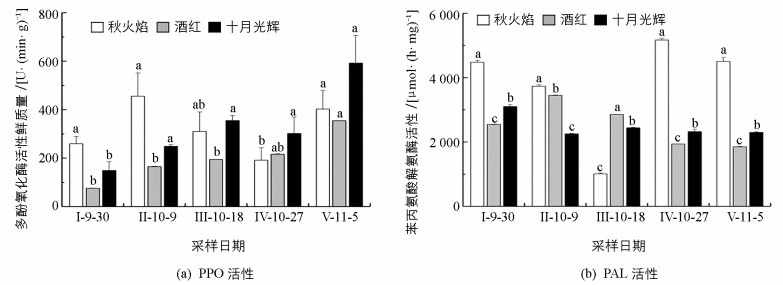
整个秋季变色期,3种北美红枫多酚氧化酶(简写为PPO)活性总体呈增高趋势,其中秋火焰PPO活性呈波浪形变化,酒红、十月光辉PPO活性则基本呈直线形变化. 11月5日与9月30日相比,秋火焰、酒红、十月光辉PPO活性(鲜质量)分别升高了144 U/(min·g)、279 U/(min·g)、443 U/(min·g). 9月30日,十月光辉PPO活性是秋火焰的0.58倍,是酒红的1.97倍;11月5日,十月光辉PPO活性是秋火焰的1.47倍,是酒红的1.67倍. 10月18日后,十月光辉PPO活性始终高于秋火焰、酒红(图 4(a)). 11月5日与9月30日相比,秋火焰苯丙氨酸解氨酶(简写为PAL)活性变化较小,酒红、十月光辉PAL活性分别下降了695 μmol/(h·mg)、802 μmol/(h·mg).整个秋季变色期,3种北美红枫PAL活性差异始终具有统计学意义,其中秋火焰PAL活性变化较为剧烈,呈“V”型,而酒红、十月光辉PAL活性变化较为平缓,且10月18日后,十月光辉PAL活性基本稳定.除10月18日秋火焰PAL活性显著低于酒红、十月光辉外,其他时期秋火焰PAL活性始终显著高于酒红、十月光辉(图 4(b)).
-
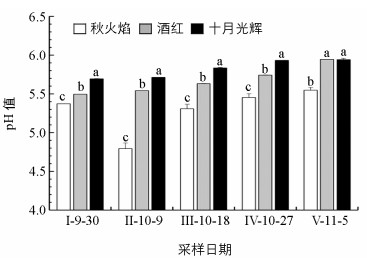
整个秋季变色期,3种北美红枫叶片pH值差异具有统计学意义,且pH值按从高到低的顺序始终为:十月光辉、酒红、秋火焰. 3种北美红枫叶片pH值始终低于土壤pH值(5.9). 11月5日与9月30日相比,秋火焰、酒红、十月光辉叶片pH值总体呈上升趋势,但分别仅上升0.17,0.45,0.25(图 5).
-
Person相关性分析结果表明(表 3),北美红枫叶片SS质量比、SP质量比、PPO活性、pH值与叶色L值、b值呈极显著负相关(p<0.01),与叶色a值呈极显著正相关(p<0.01),与叶片色素质量比及色素比呈极显著相关(p<0.01),说明此四者是影响北美红枫秋季转色的重要因子. PAL活性对北美红枫叶色L,a,b值均无显著影响,说明PAL活性不是影响北美红枫秋季转色的重要因子.
2.1. 北美红枫秋季变色期的叶色参数
2.2. 北美红枫秋季变色期的色素质量比及色素比
2.2.1. 秋季变色期的色素质量比
2.2.2. 秋季变色期的色素比
2.3. 北美红枫秋季变色期的其他生理指标
2.3.1. 秋季变色期的可溶性糖及可溶性蛋白质量比
2.3.2. 秋季变色期的相关酶活性
2.3.3. 秋季变色期的叶片pH值
2.4. 呈色因子的相关性分析
-
北美红枫适应能力强,在我国南方地区广泛引种,但北美红枫品种繁多,且表现出高度差异性,如抗寒性、生长速率、秋季着色等[19]. 3种北美红枫秋季叶色变化过程有较大差异,秋火焰的变化过程为:绿色—黄色—橙黄色;酒红为:绿色—黄色—橘黄色;十月光辉为:绿色—黄色—橙红色.与秋火焰、酒红相比,十月光辉变色较早,持续时间较长.研究报道,彩叶植物叶色变化受内外因素共同作用,是植物的应激反应之一[20].外部因素主要表现为光照、温度、湿度和土壤等方面,内部因素包括形态学、生理学和生化防御机制. 3种红枫在同一地区种植,且在形态和健康条件上相似,因此考虑叶色差异主要是由于它们的生理差异.相关性分析表明,北美红枫叶色a值与色素质量比及色素比极显著相关,此外,10月18日后,十月光辉的叶绿素、类胡萝卜素质量比始终高于秋火焰、酒红,表明北美红枫叶色变化除因色素变化外,可能还与光合作用效果有关[21].
花色素苷是一种多酚化合物,在花、叶和果实中广泛积累,在高等植物中常呈现紫色、红色等典型颜色[22].可溶性糖不仅是花色素苷合成的前体物质,同时也是一种信号物质,如与花青素合成有关的ldox基因、chs基因的表达受糖相关机制的调节[23].然而,秋火焰、酒红可溶性糖质量比远高于十月光辉,二者变色效果却远不如十月光辉,这与李梅洁等研究相一致[9],故猜测叶片内可溶性糖质量比过高,可能会抑制相关基因的表达,影响其花色素苷合成.可溶性蛋白是植物体内重要的渗透调节物质,可以帮助植物对低温和老化等不利条件做出反应.本研究发现10月9日后,十月光辉的可溶性蛋白质量比显著低于秋火焰和酒红.通过比较渗透调节物质的质量比,我们认为十月光辉比秋火焰、酒红具有更强的抗低温性.
研究表明花色素苷的合成受酶的控制[24]. PPO广泛存在于植物中,可催化酚类物质形成棕色产物,对植物色彩有一定作用[25].相关性分析表明,PPO活性和a值显著正相关,10月18日后,十月光辉PPO活性始终高于秋火焰、酒红,说明较高的PPO活性能加深北美红枫叶色,这与卓启苗对欧洲卫矛(Euonymus europaea)的研究一致[26].苯丙氨酸是花青素合成的直接前体,但关于PAL是否对花青素合成有影响一直存在争议[27].本研究中PAL活性与花色素苷无显著相关关系,说明PAL活性不是北美红枫花青素合成的关键作用因子,对其秋季叶色变化无显著影响,这与Wellman等对红毛杉(Haplopappus gracilis)的研究结果一致[28].本试验发现在北美红枫秋季变色期,其叶片pH值不断升高,且与叶色a值呈极显著正相关(p<0.01). 3种北美红枫pH值差异具有统计学意义,且十月光辉的叶片pH值始终最高,说明叶片pH值会影响北美红枫叶色转变,这与程怡等对月季花的研究结果一致[29],未来可通过适当提高土壤pH值来改善北美红枫呈色状况.本试验以3种北美红枫为材料,仅就部分生理指标探讨了北美红枫秋季叶片呈色的差异,未对其遗传因子进行分析,今后还需通过分子标记等遗传学手段开展进一步研究.
-
色素质量比、可溶性糖质量比、可溶性蛋白质量比、PPO活性、叶片pH值等是影响北美红枫转红的重要作用因子.花色素苷质量比的迅速增加、mChl/mAnt值和mCar/mAnt值的迅速降低是北美红枫转红的直接原因.北美红枫秋季变色期可溶性糖大量积累,但其过量积累可能会抑制花色素苷合成,进而影响北美红枫叶片转红. PAL活性在北美红枫秋季变色期对色素质量比及色素比并无明显影响,不是影响其叶色转变的重要影响因子.叶片pH值与北美红枫叶色a值显著相关,未来可通过调节叶片pH值人工调控北美红枫秋季变色.十月光辉在秋季变色期生长状态稳定,转红效果较好,更适合南方地区引种.