


 下载:
下载:
-
在自然界中,植物的资源在空间和时间上都是不断变化的,即生境异质性是普遍存在的[1-2].西南喀斯特地区,一方面,土层浅薄,保水能力差,岩石渗漏严重,水分亏缺成为该地区的重要特征[3].另一方面,该地区的土壤水分具有明显的时空变异性和派生性[4-6].干旱缺水和水分时空异质性严重影响该地区植物的生长、分布及繁殖,进而影响到植物群落的分布格局和稳定性,对植被演替的方向和速率产生重要影响[5, 7-8].因此,研究喀斯特地区植物对干旱及干旱条件下异质性水分生境的适应,对喀斯特地区的植被恢复和重建具有重要的理论和实践意义.
在长期的进化过程中,植物形成了一套特有的适应异质性环境的对策和机制[9-10].如在养分异质性生境中,植株将较多的根系分布在养分丰富的斑块中,即通过改变细根长度和表面积,改善根系的空间分布格局,提高总生物量生产来提高其适应性[11-13].尽管水分时空异质性普遍存在且具有重要作用,但目前研究通常只关注了克隆植物对水分空间异质性的响应[14-15],或者非克隆植物对水分时间异质性的影响[16-18],而对于非克隆植物在水分空间异质性尤其是在干旱条件下的空间异质性的响应研究极少.
复羽叶栾树(Koelreuteria bipinnata Franch.)为无患子科落叶乔木,生长迅速,树形高大,枝叶茂密,抗污染、抗病菌能力强,主要生长于石灰石风化产生的钙质土壤中,对石漠化地区典型的季节性水分亏缺环境具有很好的适应能力,在石漠化的植被恢复和重建中有极高的推广价值[19].本研究以复羽叶栾树幼苗为研究对象,通过不同土壤水分含量下水分资源的空间异质性和同质性处理,探索喀斯特地区的适生植物在干旱胁迫下对异质生境的生长和光合响应策略,以期为喀斯特地区的植被恢复提供理论依据.
全文HTML
-
实验材料为复羽叶栾树当年生幼苗,供试土壤为重庆中梁山典型黄色石灰土,土壤基本理化性质如表 1.实验在重庆市西南大学生态实验基地透明遮雨棚内进行.
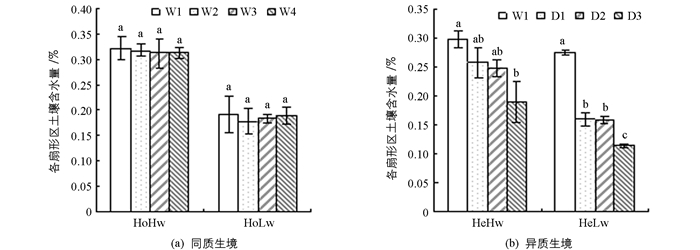
选择直径为30 cm,高24 cm的种植盆,用PVC板将花盆平均分成4个扇形区域,且中央留出直径为4 cm的小孔,以供栽植植物幼苗. 2014年6月20日,选取长势基本一致、无病无害的复羽叶栾树幼苗移栽到花盆的中央小孔,适应生长1个月后,开始水分处理.实验处理分为水分同质性生境(Ho)和水分异质性生境(He)两种;在每种生境下根据土壤的含水量设置为对照(Hw)和干旱(Lw)两种方式,根据预备实验结果,估算出对照条件下每天的浇水量为262 mL,干旱条件下每天的浇水量为126 mL.其中,同质性生境是将定量的水平均分为4份均匀地浇到花盆的4个扇形区(分为W1,W2,W3,W4区),异质性生境,则是将定量的水沿花盆壁只浇到一个固定的扇形区(W1区),其他3个区域则不浇水(另3个区域为D1,D2和D3区,其中D1和D2为缓冲区,即相邻浇水区,D3为与处理区相对的非缓冲区).每个处理3个重复.由于水分会从中央小孔向其他区域渗透,我们对每个扇形区域的土壤含水量进行了跟踪测定,以确定每种处理及每个斑块中土壤的实际含水量.
2014年8月下旬,即实验处理30 d后,停止处理.采用LI-6400便携式光合测定分析仪(LICOR,USA),选取植物顶部健康叶片,在上午10:00-11:00测定植物的净光合速率(Pn),蒸腾速率(Tr),气孔导度(Cs)等气体交换参数,并计算水分利用效率(WUE)=净光合速率(Pn)/蒸腾速率(Tr).测定完光合作用后,对植物的地上和地下部分分别进行收获,置于烘箱内,105 ℃下杀青30 min后于60 ℃烘干至恒质量,分别称量地上部分干质量(地上生物量),根干质量(根系生物量),计算总生物量,根生物量比=根系生物量/总生物量.
数据统计分析采用Microsoft Excel 2003和SPSS 21.0软件结合进行,采用双因素方差分析(Two-Way ANOVA)分析花盆内各扇形区域的土壤含水量差异以及同一水分处理条件下不同生境中光合特性指标、生物量及生物量分配的差异.
-
同质生境中,对照条件下W1,W2,W3和W4区域内的土壤含水量均在0.31~0.32之间,相当于田间持水量的88.3%,干旱条件下W1,W2,W3和W4区域内的土壤含水量均在0.18~0.19之间,相当于田间持水量的54.1%,符合Hisao[20]对水分胁迫划分标准中的轻度水分胁迫,且在两种水分处理下4个区域内土壤含水量差异无统计学意义(p>0.05).异质生境中,对照条件下,土壤含水量随W1,D1,D2和D3逐渐下降,D3区土壤含水量下降到轻度水分胁迫水平,相邻扇形区处理间土壤含水量差异无统计学意义(p>0.05),但W1区与D3区差异有统计学意义(p<0.05);干旱条件下,土壤含水量在W1,D1,D2和D3个区域内逐渐下降,D1和D2缓冲区土壤含水量下降到中度水分胁迫水平,D3区土壤含水量下降到重度水分胁迫水平,其中位置相同的D1和D2缓冲区间土壤含水量差异无统计学意义(p>0.05),其余区间差异有统计学意义(图 1).
-
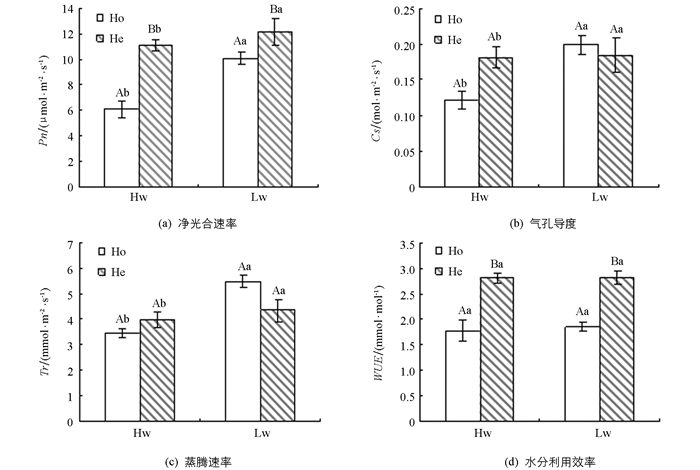
无论是在对照还是在干旱条件下,复羽叶栾树幼苗的净光合速率和水分利用效率在异质生境下均显著高于同质生境(p<0.05)(图 2a,2d),气孔导度和蒸腾速率在异质生境和同质生境下差异无统计学意义(p>0.05)(图 2b,2c).在对照条件下,净光合速率在异质生境下比同质生境下提高45.6%,在干旱条件下,净光合速率在异质生境下比同质生境下提高16.8%,而对于水分利用效率,对照的异质条件下比同质条件下提高37.3%,干旱的异质条件下比同质条件下提高34.6%.双因素方差分析表明,生境对植物的净光合速率具有极显著的影响(p<0.01),水分处理对植物的净光合速率、气孔导度和蒸腾速率的影响差异有统计学意义(p<0.05),但生境和水分处理只对蒸腾速率具有显著的交互作用(表 2).
-
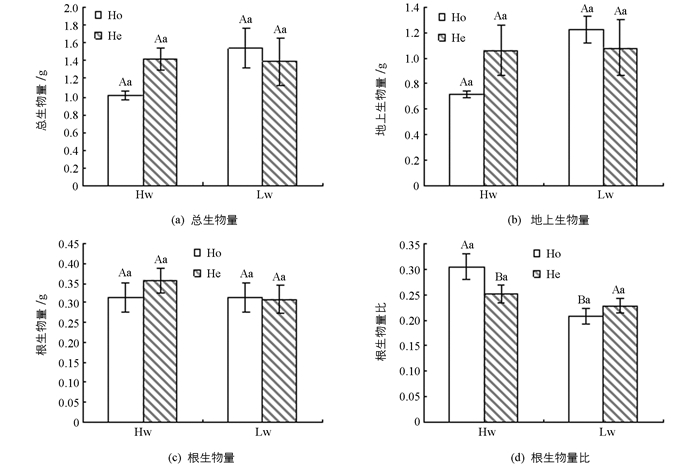
不管在对照还是干旱条件下,复羽叶栾树幼苗的总生物量、地上生物量和根生物量在同质和异质生境下差异无统计学意义(p>0.05);水分处理对生物量的积累也没有显著影响,且生境和水分没有交互作用(p>0.05)(图 3a,3b,3c,表 2).然而,对照条件下,复羽叶栾树幼苗的根生物量比在异质生境下显著低于同质生境(p<0.05),而干旱条件下,其根生物量比在异质生境下高于同质生境(p>0.05)(图 3d,表 2).水分处理对根生物量比没有显著影响,且生境和水分没有交互作用(p>0.05)(表 2).
2.1. 土壤含水量
2.2. 光合特性和水分利用效率
2.3. 生物量积累及分配
-
植物的光合作用与水分的供应和生境的差异密切相关[21].在本实验中,生境显著影响复羽叶栾树幼苗的净光合速率:不管是在对照还是干旱条件下,异质生境中复羽叶栾树幼苗的净光合速率均显著高于同质生境.与同质生境相比,异质生境中复羽叶栾树幼苗的气孔导度和蒸腾速率并未发生显著变化,但其水分利用效率显著升高,说明复羽叶栾树幼苗通过提高在异质生境中的水分利用效率来提高自身的光合速率.水分利用效率也反映了植物生产中的能量转换效率,是评价植物缺水条件下植物生长适宜程度的一个指标[22].在本实验中,异质生境中某些斑块土壤含水量明显降低,甚至达到重度干旱(低水干旱中的D3斑块),但复羽叶栾树幼苗在异质生境中能够储存大量的水分,具有更强的节水能力和更高的水分生产力,能够充分利用水资源,提高了对异质生境的适应性[14, 23-24].
通常情况下,水分减少会降低植物的光合能力[25-27].但在本研究中,不管是在同质还是异质生境下,复羽叶栾树幼苗的净光合速率在干旱条件下反而高于高水对照条件,这与万勇善等的研究结果相似,他们也发现花生在生长期内净光合速率随着水分的降低有升高的趋势[28],这可能与本实验的水分设置有关.本实验设置的干旱条件相当于田间持水量的54.1%,为轻度干旱胁迫[20].轻度干旱胁迫对有些耐旱植物不会造成伤害[29],反而有促进作用[30].在本实验中,对耐旱的复羽叶栾树来说,低水干旱条件促进了其净光合速率的增加,这也是在干旱条件下气孔导度比对照条件下有升高趋势的原因所在[19].然而,对于水分利用效率,无论在同质还是异质生境中,含水量降低并没有提高水分利用效率.一方面,本实验设置的轻度干旱条件对耐旱的复羽叶栾树没有造成干旱伤害[19];另一方面,在干旱生境中植物提高水分利用效率往往以降低光能利用效率为代价的[31],复羽叶栾树为喜光物种,在轻度的干旱条件下,也不以降低光能利用效率为代价去提高水分利用效率,从而保证了较高的光合生产.
植物生物量的积累和分配是植物对变化环境的反应[32-33].有研究表明,在异质生境下,植物的根系生长行为总是趋向于肥沃的以及水分较充足的斑块,从而提高在整个异质生境中的根系生物量[34].然而,在本实验中,异质生境中根系生物量与同质生境相比,并未发生明显改变,同时显示地上部分生物量及整个生物量均未发生显著变化.这与Stueffer等[35]研究认为白三叶的生物量在异质生境比同质生境高67%的研究结果并不一致.从“时间效应”上分析,可以看出植物的光合生理和生物量的变化在时间上具有差异,生物量是一个积累过程,其在植物生长早期对不同资源水平生境的表现形式往往比光合特征的改变时间更晚.
植物的根生物量比反应植物对资源的分配情况,其根生物量比在对照的异质生境中显著低于其对同质生境的响应,可能原因是对照的异质生境尽管造成不同斑块内含水量的差异,但含水量最低的区域仍不会对植物造成干旱伤害,植物无需向地下部分投资更多的资源,而提高地上部分的投资保证了其高的光合生产,这正是植物能够将光合作用所固定的能量进行优化分配从而提高自身在多变环境中的适合度的体现[36].干旱条件下,复羽叶栾树的根生物量比对异质生境的响应高于同质生境,说明干旱的异质生境下,植物能够通过对地下部分的投资来提高对干旱缺水的适应.
总之,复羽叶栾树通过提高水分利用效率和降低对根系资源的投资提高净光合速率,但并未提高异质生境中植物生物量的生产,可能与“时间效应”有关.