



 下载:
下载:

-
开放科学(资源服务)标志码(OSID):

-
小麦穗发芽是严重的气候灾害,是小麦生产致命性的威胁. 穗发芽灾害不发则已,一发则损失惨重. 据联合国粮农组织统计,全世界每年因穗发芽损失20.00%的小麦产量,几乎相当于我国小麦的年产量[1]. 我国黄淮麦区、长江中下游麦区曾多次发生过大面积穗发芽,西南冬麦区及东北春麦区穗发芽灾害也频繁发生. 我国有85.00%的麦区都存在穗发芽隐患[2]. 小麦穗发芽后不仅产量降低,最主要的是品质严重变劣,甚至不能食(饲)用. 连续阴雨和潮湿的气候是穗发芽灾害发生的外因,小麦品种穗发芽抗性差才是内因. 因气候环境难以控制,所以培育推广抗穗发芽的小麦品种是关键. 小麦高抗穗发芽种质资源的严重匮乏是抗穗发芽育种取得突破的主要阻碍,也是目前亟待解决的问题.
小麦的抗穗发芽育种一直是我国农业科研的重要领域,科研工作者对现有的大量小麦品种资源进行了抗穗发芽筛选鉴定,选出了一些抗性相对较好的品种作为小麦抗穗发芽育种种质资源加以利用;通过对现有材料的研究,发现了一些抗性与生理以及抗性与种皮颜色、颖壳结构等的关系[3-8],但受现有小麦品种资源遗传背景的限制并没有从根本上找到高抗穗发芽的抗源基因.
国家小麦工程技术研究中心克隆了能抑制小麦穗发芽的“反义硫氧还蛋白基因”并获得了转基因小麦材料. 经穗发芽抗性鉴定,转基因小麦种子比非转基因对照的萌动时间明显延长,对穗发芽有较强的抑制作用[9-10]. 但该材料是转基因作物,特别是目前国家严格限制在粮食作物上应用转基因的形势下,该材料在小麦抗穗发芽育种上的应用受到限制;四川农业大学利用远缘杂交技术率先将“节节麦”的高抗穗发芽基因引入小麦,创制出了小麦高抗穗发芽材料RSP[11-14],为小麦抗穗发芽育种提供了宝贵的种质资源.
在“一年两熟”种植模式的黄淮麦区,小麦收获后种植大秋作物,秋季9月底10月初大秋作物收获后,冬小麦播种耕地前,大田中往往生长有少量的刚出土不久的野生麦苗,这些野生麦苗是由小麦收获时因粗放操作遗落于大田中的麦粒,麦穗在土壤中经过了充沛降雨的夏季出土而成,其种子应该具有很强的休眠特性和抗穗发芽能力,于是我们通过收集并集中移栽的方式,对秋后野生麦苗的穗发芽抗性、种子的休眠特性、相关抗穗发芽基因的分子标记检测进行了研究,旨在为小麦抗穗发芽育种提供具有极强穗发芽抗性的种质资源.
全文HTML
-
当年的9月底10月初,秋收后冬小麦播种耕地前,对计划收集地域范围内的农田进行普查,将发现的野生麦苗(图 1)进行根部带土团移栽入观察圃,行距20 cm,株距10 cm,并进行编号,记录收集地点、移栽时间等. 本试验野生麦苗的收集地为河北省成安县商城镇赵横城村的部分农田,集中移栽地为河北省成安县商城镇赵横城村北邯郸市农业科学院生物技术研究室试验田. 将野生麦苗集中移栽入观察圃后,立即浇水保证移栽苗的成活. 并对观察圃加强管理,使移栽成活后的野生麦苗生长茁壮.
-
调查指标主要有:①株高,②抗倒性:[1级]不倒伏;[2级]轻微倒伏,植株倾斜角度A<30°;[3级]中等倒伏,30°≤A<45°;[4级]较重倒伏,45°≤A<60°;[5级]严重倒伏,A≥60°以上. ③抗病性:在大田中主要目测调查白粉病、锈病、叶枯病、赤霉病4种病害. [1等]无病(没有4种病害发生);[2等]轻微发病(有4种病害的1种或几种发生但较轻);[3等]发病较重(4种病害的1种或几种严重发生). ④越冬抗冻性:[1级]无冻害;[2级]叶尖受冻发黄;[3级]叶片冻死一半;[4级]叶片全枯;[5级]植株或大部分蘖冻死. ⑤分蘖倍数,⑥有效分蘖率,⑦穗粒数,⑧千粒质量等.
-
穗发芽抗性测定参照中华人民共和国农业行业标准《小麦抗穗发芽性检测方法》(NY/T 1739-2009)[15]进行. 在小麦蜡熟后期,从田间采取野生单株和对照“泰山1号”植株的主茎穗各10穗,从穗下茎15~20 cm处剪取. 将剪取的每单株的10个麦穗随机分成2组,每组5穗. 先用自来水浸泡4 h,然后用0.10%的次氯酸钠溶液消毒5 min,再用无菌水反复冲洗. 将麦穗插在泡沫塑料板上,放在人工气候箱中,每天模拟降雨喷水2次,温度为22 ℃,相对湿度为100.00%,培养96 h后立即放在60 ℃的烘箱中烘干. 然后手工剥粒调查统计每组麦穗的发芽粒数(以种子籽粒胚部种皮破裂为发芽标准)和总粒数.
计算穗发芽率(X)
相对穗发芽指数(I)
式中,N发芽为试验穗发芽粒数,N总为试验穗总粒数,X1为待测样品的穗发芽率,X2为对照品种的穗发芽率.
进一步对抗穗发芽级别进行评价(I=0为极抗,I<0.05为高抗,0.05≤I<0.20为抗, 0.20≤I<0.40为中抗,0.40≤I<0.60为感,I≥0.60为高感).
-
根据试验情况,把同时符合相对穗发芽指数I<0.05,株高H≤75 cm,抗倒性为[1级],抗病性为[1等],越冬抗冻性为[1级],分蘖倍数R≥20,有效分蘖率ER≥80.00%,穗粒数N≥35粒,千粒质量W≥45 g几项标准的野生麦苗作为种质资源利用,其余的淘汰.
-
将符合本文1.4标准入选的野生麦苗抗穗发芽单株全部种子按株系种植,行距20 cm,株距10 cm,单穴单粒播种. 次年在小麦蜡熟后期,对入选的野生麦苗抗穗发芽种质系进行穗发芽抗性鉴定. 对麦穗的处理方法和培养条件与本文1.3部分相同,不同的是:培养时间为15 d. 从开始培养每间隔24 h调查统计1次各种质系麦穗籽粒的发芽数量,以籽粒生出种根或长出麦芽为发芽标准. 每天调查后去除已经生根、发芽的麦粒,剩下未发芽的籽粒继续培养直到15 d试验结束. 计算出每天的穗生根发芽率并制作“时间-穗生根发芽率”曲线.
-
从小麦蜡熟后期开始,在25 ℃的室温下,对各个野生麦苗种质系的种子,每间隔3 d进行1次发芽试验,96 h后调查发芽率,当种质系种子的发芽率达到80.00%以上时可认为种子休眠期结束,即种质系种子发芽率达到80.00%以前的时间为该种质系种子的最长休眠期.
-
Vp1B3(F:5′TGCTCCTTTCCCAATTGG3′;R:5′ACCCTCCTGCAGCTCATTG3′)是内蒙古农牧大学杨燕等[16]开发的与小麦某些抗穗发芽基因紧密相关的STS分子标记,利用Vp1B3在抗穗发芽小麦品种中能扩增出845 bp和569 bp两种片段类型,而在感穗发芽品种中仅扩增出652 bp的片段类型. TaDFRBb(F:5′GAAATGCGGGGTGACTAGGACTCT3v;R:5′TATTTCCGTGGCTATTGATTGTTTC3′)是我国开发的与小麦红皮基因紧密相关的抗穗发芽分子标记,在红皮抗穗发芽小麦品种中能扩增出534bp的A型片段和526 bp的B型片段[17-19].
采用CTAB法提取抗穗发芽种质系的基因组DNA,Vp1B3和TaDFRBb引物由上海生工合成. 构建25 μL的PCR反应体系[10×Buffer 2.50 μL+dNTP Mix(2.50 mmol/L)2 μL+上、下游引物(20 μmol/L)各0.50 μL+Taq酶(5U/μL)0.25 μL+模板DNA 1 μL+ddH2O 18.25 μL].
PCR反应程序:94 ℃预变性5 min→94 ℃变性1 min→退火1 min(Vp1B3 60 ℃;TaDFRBb 59 ℃→72 ℃延伸1 min(以前做35次循环)→72 ℃延伸10 min,PCR反应结束. 用1%的琼脂糖凝胶电泳检测PCR反应产物,EB染色后用凝胶成像拍照.
1.1. 野生麦苗的收集与集中移栽
1.2. 集中移栽的野生麦苗植株农艺性状调查
1.3. 集中移栽野生麦苗麦穗的相对穗发芽指数测定
1.4. 野生麦苗抗穗发芽种质系的入选标准
1.5. 野生麦苗抗穗发芽种质系穗发芽抗性测定
1.6. 野生麦苗抗穗发芽种质系种子的休眠期测定
1.7. 抗穗发芽基因相关STS分子标记检测
-
由表 1知,当年的9月底10月初生长于田间的野生麦苗,根部带土移栽很容易成活,只要移栽后及时浇水,移栽成活率为100.00%.
-
通过对稳定遗传的118个野生麦苗单株的主要农艺性状指标进行调查,结果显示(表 2),野生麦苗植株都比较高,有65.30%的野生麦苗单株的株高在100 cm以上,有79.70%的野生麦苗单株的株高在80 cm以上,20.40%的野生麦苗单株的株高低于80 cm,只有6.80%的野生麦苗单株的株高不高于75 cm. 野生麦苗单株的抗倒性大部分较差,有65.30%的野生麦苗单株发生中等及以上倒伏,21.20%的野生麦苗单株倒伏较轻,只有13.60%的野生麦苗单株不发生倒伏. 野生麦苗植株的抗病性(白粉病、锈病、叶枯病、赤霉病)普遍较好,在大田环境中80.50%的野生麦苗植株无病害发生,只有19.50%的野生麦苗植株轻微发病. 野生麦苗植株的越冬抗冻性都较好,越冬冻害没有高于[2级]的单株,55.90%的野生麦苗植株越冬冻害为[1级]. 野生麦苗植株的分蘖倍数和有效分蘖率都较高,100.00%的野生麦苗植株的分蘖倍数都大于20倍,且有效分蘖率都不低于80.00%. 野生麦苗单株的穗粒数较多,100.00%的野生麦苗单株穗粒数都不低于35粒,但是野生麦苗单株的千粒质量较小,在行距20 cm,株距10 cm,单穴单粒种植的情况下,78.80%的野生麦苗单株的千粒质量低于45 g,只有21.20%的野生麦苗单株的千粒质量不低于45 g.
-
分析表 3对118个稳定遗传野生麦苗单株(系)的穗发芽抗性测定数据可知,野生麦苗的穗发芽抗性普遍较好,有19.50%的野生麦苗单株的相对穗发芽指数为0,有78.80%的野生麦苗单株的穗发芽抗性都能达到高抗水平,只有17.80%和3.40%野生麦苗单株的穗发芽抗性为抗和中抗.
-
根据本试验制定的野生麦苗抗穗发芽种质系的入选标准,符合条件的野生麦苗抗穗发芽种质系集合{野生麦苗抗穗发芽种质系}={相对穗发芽指数I<0.05}∩{株高H≤75 cm}∩{抗倒性为[1级]}∩{抗病性为[1等]}∩{越冬抗冻性为[1级]}∩{分蘖倍数R≥20}∩{有效分蘖率ER≥80.00%}∩{穗粒数N≥35粒}∩{千粒质量W≥45 g}={21,46,63,106},图 2为穗发芽试验照片.
-
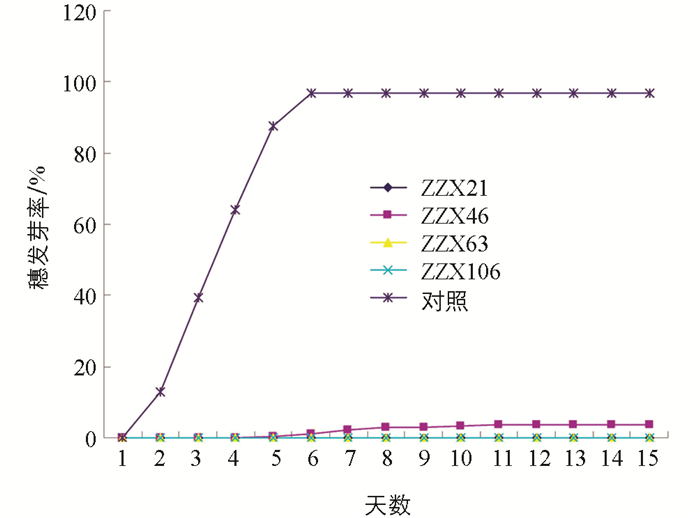
由表 4和图 3可知,4个野生麦苗抗穗发芽种质系进行1~15 d的模拟穗发芽试验时,对照“泰山1号”(CK)第2 d就开始在穗上发芽,第3 d时穗发芽的粒数最多,第6 d时能发芽的麦粒全部在穗上发芽,累计穗发芽粒数为376粒,穗发芽率达到了96.91%,最后仅剩下12粒由于种子质量的原因未能发芽. 抗穗发芽种质系ZZX21,ZZX63,ZZX106经15 d的模拟穗发芽试验穗发芽率皆为0,而且所有的麦粒状况良好均未发生霉烂. ZZX46在模拟穗发芽试验的第5~12 d中,每天有少量的麦粒在穗上生根或发芽,第12 d时累计穗发芽粒数最多为15粒,穗发芽率为3.79%,模拟试验15 d时未发芽的麦粒状况良好均未发生霉烂. 经15 d模拟穗发芽试验,ZZX21,ZZX46,ZZX63,ZZX106 4个野生麦苗抗穗发芽种质系的相对穗发芽指数分别为0,0.039,0,0,均小于0.05达到高抗级别.
-
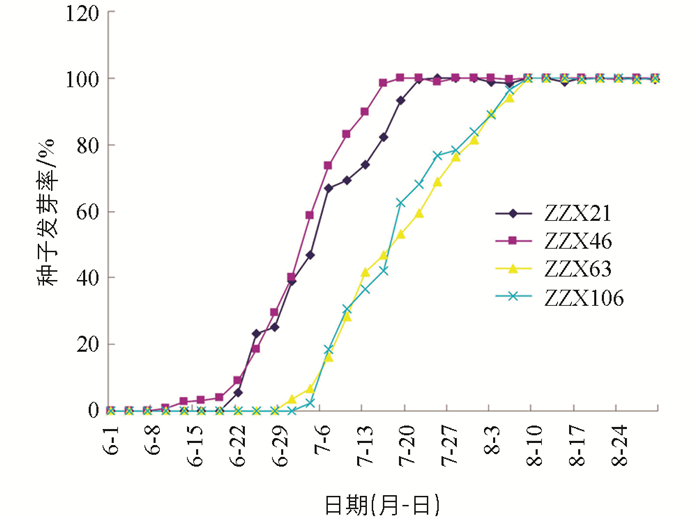
由表 5和图 4可知,抗穗发芽种质系ZZX21和ZZX46从种子生理成熟(蜡熟期)6月1日开始到7月16日和7月10日时种子发芽率才达到80.00%以上,休眠时间分别为46 d和40 d. 抗穗发芽种质系ZZX63和ZZX106从种子生理成熟(蜡熟期)6月1日开始到7月31日时种子发芽率才达到80.00%以上,休眠时间为61 d. 抗穗发芽种质系ZZX21,ZZX46,ZZX63,ZZX106种子都有较长的种子休眠期,在大田种子生理成熟后,对连续阴雨的气候环境引起的小麦穗发芽有很好的抗性. 其最长休眠期通常能在40~60 d结束,不影响10月初的冬小麦大田播种出苗.
-
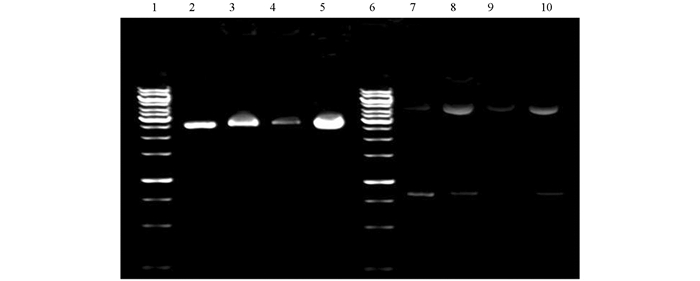
由图 5抗穗发芽分子标记检测结果可知,在抗穗发芽种质系ZZX21,ZZX46,ZZX63,ZZX106基因组DNA中未检测出Vp1B3和TaDFRBb的阳性条带,这说明野生麦苗抗穗发芽种质系的相关抗穗发芽基因不属于和Vp1B3或TaDFRBb紧密连锁的已知抗穗发芽基因.
2.1. 秋后野生麦苗收集移栽统计
2.2. 野生麦苗植株主要农艺性状调查结果
2.3. 野生麦苗株系相对穗发芽指数的测定
2.4. 野生麦苗抗穗发芽种质系的筛选结果
2.5. 野生麦苗抗穗发芽种质系的穗发芽抗性测定结果(培养15 d)
2.6. 野生麦苗抗穗发芽种质系种子的休眠期测定结果
2.7. 抗穗发芽基因相关STS分子标记检测结果
-
① “一年两熟”制的黄淮麦区,秋后9月底10月初,冬小麦播种耕地前,大田中刚出土不久的野生麦苗,具有很强的抗穗发芽能力,通过集中移栽,相对穗发芽指数测定,有78.80%的秋后野生麦苗穗发芽抗性达到高抗水平,19.50%的秋后野生麦苗相对穗发芽指数为0,穗发芽抗性达到极抗水平.
② 本研究通过集中移栽、综合农艺性状调查、模拟穗发芽试验等选出了4个野生麦苗抗穗发芽种质系ZZX21,ZZX46,ZZX63,ZZX106,其相对穗发芽指数分别为0,0.039,0,0,均达到了极抗穗发芽水平. 其种子的休眠期较长但不影响10月初的冬小麦播种出苗.
③ Vp1B3和TaDFRBb是当前开发最成功也是应用较广泛的小麦抗穗发芽基因分子标记. 通过这两种分子标记检测发现,ZZX21,ZZX46,ZZX63,ZZX106种质系的抗穗发芽基因不属于与Vp1B3或TaDFRBb紧密连锁的抗穗发芽基因,而是新的基因类型.
④ 小麦的穗发芽抗性是由多基因控制的数量性状,在这些控制基因中有个别主效基因起着主导作用[20]. 能够抑制α-淀粉酶活性的基因(如α-淀粉酶抑制蛋白基因等)对小麦穗发芽抗性起着主效基因的主导作用,因为在小麦穗发芽过程中α-淀粉酶对淀粉的水解起着关键的作用. 张海峰等[21]的研究表明,α-淀粉酶的动态变化与穗发芽率的变化完全一致,呈极显著正相关. 因此采取各种手段抑制小麦种子内源α-淀粉酶的活性是解决小麦穗发芽问题的关键.
⑤ 秋后出土的野生麦苗是由当年5至6月份小麦收获时因粗放操作遗落于大田中的部分麦粒(穗)在土壤中经过了充沛降雨的夏季,直到9月底10月初才出土成苗,其种子具有很强的休眠特性和抗穗发芽能力. 这充分说明了在自然界小麦生产庞大而复杂的群体中存在着抗穗发芽基因型. 通过对秋后野生麦苗的收集与集中移栽,可以选出高抗穗发芽且综合农艺性状较好的抗穗发芽种质资源系,为小麦抗穗发芽育种提供了宝贵的新种质资源.