


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
中国是世界最大的柑橘生产国,柑橘产业已发展为中国南方地区农业支柱产业之一[1-2]。三峡库区作为全国第四大柑橘主产区,截至2022年,区内柑橘种植面积已近388万hm2,产量约390万t[3]。柑橘园作为一种提供较高经济收益的典型农业生态系统易受到人为干扰,频繁的人为干扰破坏土壤团聚体稳定性,增加柑橘园水土流失风险,导致土壤质量和生产能力下降[4]。绿肥覆盖种植能减少对柑橘园的人为干扰,有效增加土壤有机质含量,提高土壤团聚体稳定性,减少水土流失,改善土壤结构,有效提升土壤质量[5-6]。因此,在三峡库区柑橘园开展不同绿肥品种对土壤团聚体稳定性影响的研究,对选择适宜的果园绿肥覆盖品种,促进柑橘产业可持续发展意义重大。
团聚体是土壤结构的基本单元,是反映土壤质量及其生产功能的重要指标[7-8],在改善土壤结构、减少水土流失、提高土壤质量、防止土壤退化等方面有着重要作用[9]。团聚体的形成和稳定受土壤质地、有机质含量、地表植被特性等因素的影响。在团聚体的形成过程中,黏粒是微团聚体(<0.25 mm)的重要组成部分,能促进微团聚体的形成[10],有机质作为重要胶结物质[11],能将微团聚体和较大的粉粒、沙粒黏合形成大团聚体(>0.25 mm),使团聚体的结构更为稳定[12-14]。三峡库区紫色土分布广泛,其成土较快,结构疏松,黏粒和有机质含量较低,团聚体稳定性较差,致使库区柑橘园极易产生严重的水土流失和土壤退化[15]。柑橘园绿肥覆盖种植后能显著提升表层土壤有机质含量,促进团聚体的形成和稳定,减少水土流失,提升土壤生产功能[16]。绿肥植株体和其凋落物增加了地上生物量归还,促进了土壤中有机质的形成和积累,是绿肥提升土壤有机质含量的重要途径[17-18]。有机质含量的提升使土壤胶结作用增强,提升了大团聚体比例和稳定性,提高了柑橘园土壤的抗侵蚀性[19]。绿肥根系的生长能压实包裹土壤颗粒,根系分泌物能促进土壤中有机质的形成和累积,并能促进土壤中小型动物的生长[20],进而从物理、化学和生物等途径促使微团聚体的形成,提升大团聚体比例,促进团聚体的合理分布和稳定[21]。绿肥生长较快、覆盖好,能显著增加地表覆盖度,从而遮挡降雨,降低雨滴对团聚体的击溅分散作用,绿肥植株茎秆能减缓地表径流流速,减小地表径流对土壤颗粒和团聚体的剥蚀搬运能力,维持团聚体的稳定[22]。由此可见,绿肥的生长特性直接影响团聚体的分布和稳定性。然而,不同绿肥品种在相同季节,相同绿肥品种在不同季节的生长特性均存在显著差异,对土壤团聚体分布和稳定性的影响也不尽相同。重庆降雨量大且时间分布不均,由于降雨是土壤侵蚀的主要驱动因素之一[23],致使柑橘园在不同季节的侵蚀状况存在较大差异,因此需要选择适宜的绿肥品种,以匹配三峡库区柑橘园侵蚀状况的季节动态变化特征。虽然绿肥覆盖种植对团聚体分布和稳定性的影响效果已得到证实[24],但季节动态变化下绿肥覆盖种植对三峡库区柑橘园土壤团聚体分布和稳定性的影响尚不清楚。
鉴于此,本研究通过在三峡库区忠县橘园进行为期一年的定位试验,以白花三叶草(Trifolium repens L.)、早熟禾(Poa annua L.)、毛叶苕子(Viciavillosa Roth)3种不同绿肥覆盖下的柑橘园系统为研究对象,以传统清耕处理作为对照,深入分析不同绿肥覆盖处理下柑橘园土壤理化性质、植被特性和土壤团聚体稳定性的季节变化特征,明确不同绿肥在季节动态变化条件下对团聚体分布和稳定性的影响及主控因素,以期为选择适宜的果园绿肥覆盖品种,促进柑橘产业可持续发展提供理论支撑。
全文HTML
-
研究区位于重庆市忠县新立镇三岔湾柑橘园(30°15′57″N,107°37′53″E)。忠县位于重庆市中部,地处三峡库区中心,属于典型丘陵地貌,主要土壤类型为紫色土。该区域平均海拔462.75 m,为亚热带东南季风区山地气候,四季分明,雨量充沛,日照充足。年平均气温17 ℃,年平均降水量1 279 mm,日照时数1 300~1 400 h,日照率29%,无霜期341 d。
研究区柑橘园内的柑橘树均为2006年种植,2018年后陆续嫁接,并种植生草,果树在2-3月施基肥(高氮复合肥)、6-7月施追肥(复合肥)、9-10月施追肥(高钾复合肥),12月套袋,次年1月采果后施基肥(柑橘皮渣有机肥),管理方式统一。园内覆盖草种主要以禾本科植物早熟禾、豆科植物白花三叶草、毛叶苕子为主。试验设置白花三叶草、早熟禾、毛叶苕子3种不同绿肥覆盖处理,以传统清耕处理作为对照,样地基本情况如表 1所示。其中,清耕处理样地全年以自然生草为主,每个月进行一次割草。绿肥覆盖样地在植被生长过程中不额外施肥,撒播量以全园覆盖为准。白花三叶草、早熟禾为多年生绿肥,不额外播种。毛叶苕子在秋季播种,5月时进入结荚枯萎期。
-
在每个样地分别选取3个1 m×1 m且具有代表性的样方,收集植物样本于塑封袋中带回实验室烘干,称取干质量用于计算地上生物量;用500 cm3(直径6.37 cm、高度10 cm)的环刀取样,用于测定0~10 cm范围土层的植被根重密度;每个样方内用100 cm3(直径5.20 cm、高度5.15 cm)的环刀取样,用于测定0~10 cm范围土层的土壤容质量。不同样地均采用五点取样法采集0~10 cm范围土层的散土样品,每个样地采集2 kg散土,散土采集后放入塑封袋带回实验室,去除石砾和根系后放在室外阴凉通风处自然风干,用于测定土壤机械组成、土壤有机碳、土壤团聚体。
-
土壤容质量采用环刀法进行测定,将采回的100 cm3环刀样品浸泡至饱和,称取饱和质量,然后放置于烘箱内105 ℃烘干24 h,称取干质量,计算容质量及孔隙度[25]。土壤机械组成采用激光粒度仪(Mastersizer 3000)进行测定[26]。土壤有机碳采用重铬酸钾外加热法进行测定[27]。
水稳性团聚体采用Yoder湿筛法测定[28],湿筛后取出筛网中各级团聚体土样冲洗放置于铝盒中,在70 ℃条件下烘干,称取各粒级团聚体质量。
-
1) 植被覆盖度:在样地内随机或系统布置若干个样方,测量样方内植被覆盖的面积,计算出每个样方的植被覆盖度,然后取平均值作为整体覆盖度[29]。
2) 地上生物量:将各样方内采回的植物样品60 ℃烘干,称取并记录干质量(g)[30]。
3) 根重密度:将500 cm3环刀土样用筛网和水流小心清洗,分离出土壤中的根系。随后将分离出的根系放置在烘干箱中,60 ℃烘干至恒质量后,记录其干质量(mg)[31]。
-
采用平均重量直径(Mean Weight Diameter,MWD)、几何平均直径(Geometrical Mean Diameter,GMD)[32]和分形维数(D)[33]来评价土壤团聚体的稳定性。
土壤团聚体MWD的计算公式为:
式中:Xi为i粒级团聚体的平均直径(mm);Wi为i粒级团聚体的比例(%)。
土壤团聚体GMD的计算公式为:
土壤团聚体D值的计算公式为:
式中:Xmax为最大粒级团聚体的平均直径(mm);W(i≤Xi)为粒级小于Xi的团聚体质量(g);W0为土壤各粒级团聚体质量总和(g)。
-
采用Microsoft Excel 2010软件对试验数据进行整理和计算,采用SPSS 22.0软件对数据进行统计分析。采用单因素方差分析(One-way ANOVA)检验每个月不同处理下各个指标的差异,显著性水平为(p<0.05);利用Pearson相关分析设定置信区间95%,对|r|>0.6的强相关性进行FDR校正,对土壤理化性质、植被特性、土壤团聚体组成分布及稳定性等指标相关性进行分析;主控因素识别通过RDA分析(CANOCO 5.0)完成,使用Origin 2024软件进行制图。
1.1. 研究区概况
1.2. 试验设计与样品采集
1.3. 样品测试分析
1.3.1. 土壤理化性质测定
1.3.2. 植被特性测定
1.3.3. 土壤团聚体评价
1.4. 数据处理
-
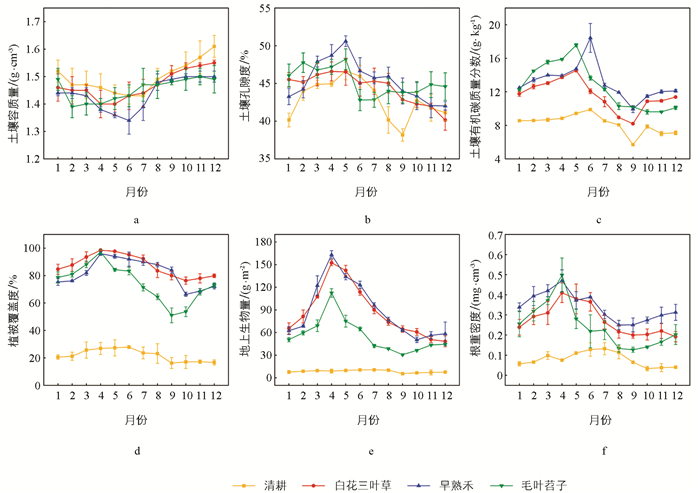
如图 1所示,同一月份不同品种绿肥种植条件下,土壤理化性质和植被特性存在显著差异。不同品种绿肥种植条件下,土壤理化性质表现为:清耕处理下,土壤容质量较高,在研究期内土壤较为紧实,孔隙度较低。绿肥覆盖下,土壤容质量在整个生长季呈上升趋势,但在4-6月较低;孔隙度在整个生长季呈下降趋势,白花三叶草和早熟禾覆盖下4-6月孔隙度较高,早熟禾在5月达到最高值50.60%,毛叶苕子覆盖下孔隙度在1-5月较高,枯萎后下降。
清耕处理下,土壤有机碳质量分数全年呈较低水平。而在绿肥覆盖处理下,土壤有机碳质量分数在整个生长季呈下降趋势,其中上半年整体较高。从具体表现来看,毛叶苕子处理的有机碳质量分数在1-5月高于其他两种绿肥覆盖处理,早熟禾处理则在6月达到最高值18.43 g/kg。
不同品种绿肥种植条件下,植被生长特性表现为:清耕处理下,植被覆盖度、地上生物量、根重密度在研究期内中呈较低水平。绿肥覆盖处理下,植被特性呈现出明显的季节性波动,3-6月植被覆盖度、地上生物量、根重密度较高,随后呈下降趋势。
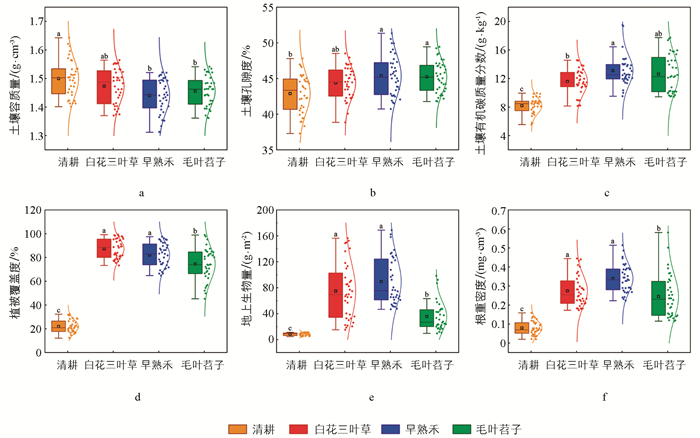
如图 2所示,在研究期内不同品种绿肥种植条件下,土壤理化性质和植被特性存在显著差异。不同品种绿肥种植条件下,土壤理化性质表现为:清耕处理下,土壤容质量显著高于绿肥覆盖处理,而土壤孔隙度和土壤有机碳质量分数显著低于绿肥覆盖处理,其中早熟禾、毛叶苕子覆盖下,土壤容质量和孔隙度无显著差异。但在毛叶苕子覆盖下,土壤有机碳质量分数的离散度较大,显示出土壤有机碳的分布存在较大差异。
不同品种绿肥种植条件下,植被生长特性表现为:清耕处理下,植被覆盖度、地上生物量、根重密度均显著低于绿肥覆盖处理,与清耕处理相比,根重密度分别显著提升70.97%、76.55%、67.47%。
-
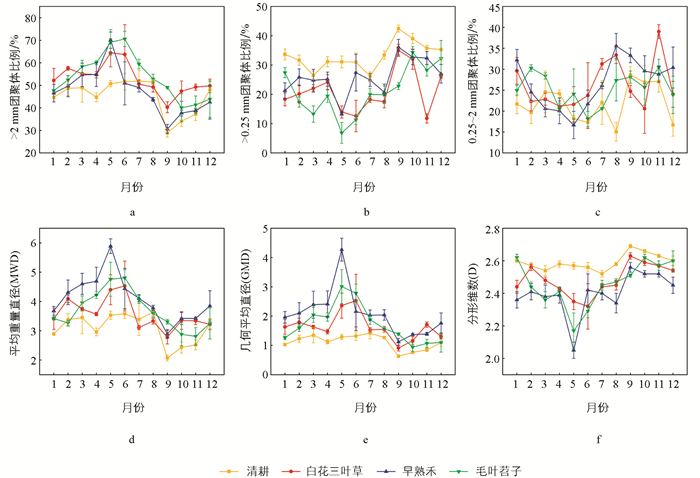
如图 3所示,同一月份不同品种绿肥种植条件下,土壤团聚体分布和稳定性存在显著差异。不同品种绿肥种植条件下,土壤团聚体分布表现为:清耕处理下,>2 mm团聚体、0.25~2 mm团聚体比例全年呈较低水平,<0.25 mm团聚体比例则较高。绿肥覆盖处理下,各级团聚体比例具有显著的季节性波动趋势。4-6月,>2 mm团聚体、0.25~2 mm团聚体比例较高,随后呈下降趋势,10-12月回升,<0.25 mm团聚体比例反之。
不同品种绿肥种植条件下,土壤团聚体稳定性表现为:清耕处理下,MWD、GMD全年呈较低水平,D值则较高。绿肥覆盖处理下,各个团聚体稳定性指标具有显著的季节性波动趋势,MWD、GMD在4-6月较高,随后呈下降趋势,D值在4-6月较低,随后呈上升趋势。值得注意的是,在早熟禾覆盖下,MWD、GMD在5月时分别达到最高值5.89、4.27,D值达到最低值2.05。
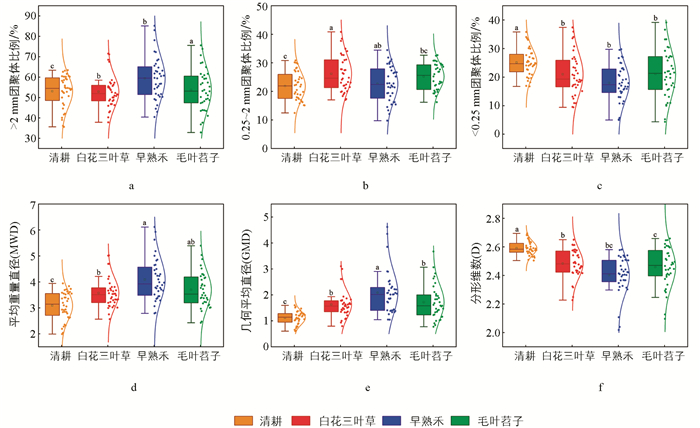
如图 4所示,在研究期内不同品种绿肥种植条件下,土壤团聚体分布和稳定性存在显著差异。绿肥覆盖处理下,>2 mm团聚体比例显著高于清耕处理,其中早熟禾覆盖下>2 mm团聚体比例显著高于白花三叶草、毛叶苕子覆盖。而在白花三叶草覆盖下0.25~2 mm团聚体比例较高,显著高于毛叶苕子和清耕处理。清耕处理下<0.25 mm团聚体比例显著高于白花三叶草、早熟禾覆盖,而毛叶苕子覆盖与清耕处理无显著差异。
清耕处理下,MWD、GMD均显著低于3种绿肥覆盖处理,而D值显著高于有绿肥覆盖处理的样地,其中早熟禾覆盖下,其GMD显著高于白花三叶草、毛叶苕子覆盖。
-
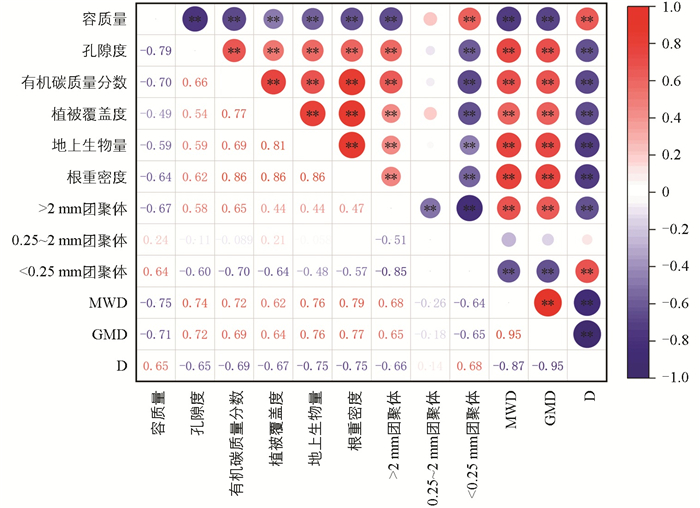
如图 5所示,Pearson相关分析结果表明,MWD、GMD分别与孔隙度、有机碳质量分数、植被覆盖度、地上生物量、根重密度、>2 mm团聚体比例呈极显著正相关(p<0.01),与<0.25 mm团聚体比例呈极显著负相关(p<0.01)。分形维数(D)与<0.25 mm团聚体比例呈极显著正相关(p<0.01),与孔隙度、有机碳质量分数、植被覆盖度、地上生物量、根重密度、>2 mm团聚体比例呈极显著负相关(p<0.01)。
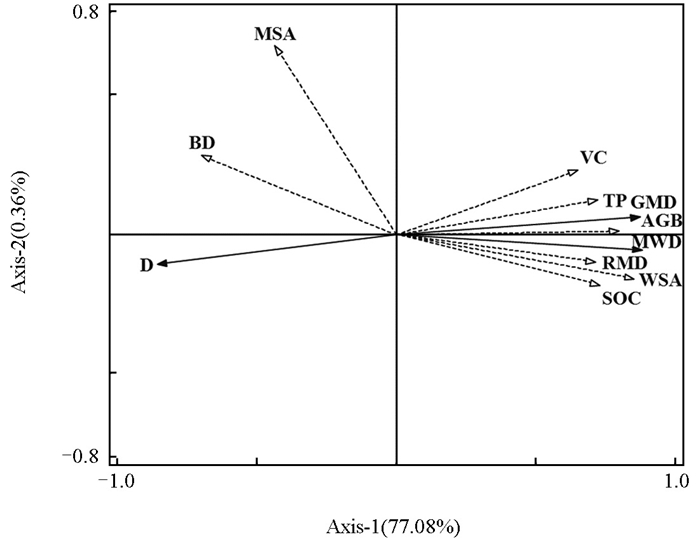
综上所述,土壤团聚体粒径分布和稳定性受孔隙度、有机碳质量分数、植被覆盖度、地上生物量、根重密度等因素的共同影响,因此采用冗余分析法进一步探究导致不同绿肥覆盖对土壤团聚体分布和稳定性存在差异的主控因素。结果表明,本研究测定的土壤性质和植被特性可以解释不同植被类型土壤团聚体稳定性77.44%的差异,其中第一轴和第二轴的解释率分别为77.08%和0.36%(图 6)。所有影响因素中,不同绿肥覆盖下,土壤团聚体稳定性受主控因子>2 mm团聚体比例影响,其解释率为56.10%(相对贡献率72.40%),显著性:p=0.002。大团聚体(>2 mm)通过机械保护作用增强抗剪强度,是维持土壤结构稳定的关键。地上生物量(AGB)解释率为17.20%(相对贡献率22.20%),显著性:p=0.002。孔隙度(TP)解释率为2.50%(相对贡献率3.20%),显著性:p=0.002(表 2)。
2.1. 不同绿肥覆盖下土壤性质和绿肥植被特性的季节变化特征和差异
2.2. 不同绿肥覆盖下土壤团聚体分布和稳定性的季节变化特征和差异
2.3. 土壤团聚体分布和稳定性的主要影响因素分析
-
季节性水热条件的改变,影响着土壤理化性质和植被特性,进而调控土壤有机碳的含量和团聚体的形成。清耕处理下,土壤团聚体稳定性在研究期都呈现较低水平,这是因为清耕处理不仅破坏了地表植被覆盖,还破坏了土壤结构,导致大团聚体破碎,微团聚体比例增加。值得注意的是,在团聚体粒径分布分析中,粒径为<0.25 mm的团聚体变化幅度相对较大,这可能是因为在发生土壤侵蚀和人为干预时,较大粒径的土壤团聚体更易受到外力破坏成为微团聚体,而粒径为0.25~2 mm的团聚体变化幅度则相对稳定[34]。
在绿肥覆盖下,春季良好的水热条件促进植物的生长发育,表现为地上生物量增加,根系伸长生长,土壤孔隙增大。地上生物量是土壤有机碳持续、间接的来源,作为植物在其生命周期内不断积累的有机物质,根、茎、叶随着季节变化自然脱落为枯落物,进入土壤中并经过微生物分解成为土壤有机碳[35-36]。随时间推移,植物根系向土壤源源不断地输送分泌物(如有机酸、可溶性糖)等根际有机质,能够促进土壤有机碳的积累[37],另外,植物根系通过物理穿插、缠绕作用又将土壤颗粒固结在一起[38-39],粒径>2 mm的土壤团聚体比例上升,团聚体稳定性也得到提高。
在植物生长旺盛时,植被覆盖度和地上生物量较高,根系活动使得分泌物和有机质输入达到高峰,土壤结构得到有效改善。在夏季6-8月,伴随高温而来的还有热胁迫,若土壤水分供应不足,会影响地上植被的蒸腾作用,抑制根系的生长,降低根际生物的活性[40-41],从而调控土壤有机碳的含量和团聚体的形成。9-11月,气温逐渐降低,降雨减少,植物生长减缓,根系活动减弱,已形成的土壤团聚体开始进入稳定状态,这时候植被覆盖度降低,地上生物量减少,若缺乏新的有机质投入,部分团聚体可能逐渐破裂或变小,有机碳也会被暴露、分解。
不同绿肥品种对土壤团聚体稳定性的影响存在差异。本研究表明,早熟禾在改善土壤结构的效果上优于白花三叶草和毛叶苕子。在以往的研究中,与豆科绿肥相比,禾本科绿肥虽没有固氮的作用,但其碳氮比较高,提高土壤有机质的效果明显[42],这与本研究结果一致。不同植物类型具有不同的根系类型(须根系和直根系),这可能是造成不同绿肥品种对土壤团聚体稳定性影响不同的主要原因之一。白花三叶草和毛叶苕子作为直根系豆科植物,根系发达,能够穿插在土壤中形成较多的孔隙,主根随土层深度增加逐渐变细,根系生物量较高[43];早熟禾作为须根系禾本科植物,细根发达,能在土壤中穿插并与其充分接触,整体分布较为均匀[44]。赵富王等[45]研究表明,须根系植物能够显著改善0~20 cm表层土土壤结构稳定性。相比于直根系植物,须根能够在0~20 cm表层土形成密集的根网,有效增加土壤颗粒之间的物理黏合,防止颗粒分散,从而提高土壤团聚体的稳定性。同时,细根生命周期较短,常年不断更新,这些死亡的根系残体在土壤中分解,成为土壤有机质的重要来源[46]。
-
相关性分析表明,土壤大团聚体比例和团聚体稳定性与土壤孔隙度、有机碳质量分数、植被覆盖度、地上生物量、根重密度等呈极显著相关。长期清耕会导致土壤退化,压实土壤,增加土壤容质量,降低土壤孔隙度,影响果园产量。植被覆盖能够降低容质量,增加土壤孔隙度和土壤黏粒含量[47]。良好的土壤孔隙结构有利于水分和空气的流通,为植物根系和微生物的活动创造适宜的环境,从而提高土壤团聚体的稳定性。
土壤有机碳是土壤团聚体形成的关键黏合剂,有机碳通过其胶质性增强了土壤颗粒的黏结力,它将微团聚体黏结形成大团聚体,同时植物根系的挤压和微生物菌丝的缠绕也是大团聚体形成的重要途径[48]。薛欣欣等[49]研究发现,在海南橡胶园种植豆科绿肥后,0~20cm土层SOC显著提升19.8%,并且提高了MWD和GMD。薛欣欣等[49]研究表明,有机碳质量分数较高时,更易促进大团聚体形成,改善土壤的水稳性。另外,团聚体通过物理保护作用将有机碳包裹在复合体中,使其免受土壤酶的攻击和土壤微生物的矿化分解[50],减少了有机碳的损失。Zhong等[51]研究发现,粒径>2 mm的团聚体对有机碳的保护性更强。
地上生物量通常作为土壤有机碳源的储存库,当植物凋零时,残体进入土壤,经过微生物分解成为有机碳[52]。植被覆盖能够显著增加根重密度和分泌物来促进土壤有机碳的积累,从而提高大团聚体比例和团聚体稳定性。有研究指出,土壤碳库中有30%~80%的有机碳是通过细根的快速周转和分解提供的[53],且根系分泌物对土壤有机碳的贡献度比同等质量的地上枯落物高出约5倍[54],尤其是植物细根数量越多,土壤碳的输入量也会越高[55]。
-
1) 绿肥覆盖下土壤性质和植被特性存在显著的季节变化规律,在4-6月时绿肥对土壤结构改良的效果最佳。同时,与清耕相比,绿肥覆盖显著地增加土壤孔隙度、根重密度和有机碳质量分数,其中早熟禾效果最佳。
2) 绿肥覆盖下土壤团聚体分布和稳定性存在显著的季节变化规律,在4-6月时大团聚体比例最高,稳定性最强。同时,与清耕相比,绿肥覆盖处理下的>2 mm团聚体比例、MWD和GMD显著增加,<0.25 mm团聚体比例与D值显著降低,其中早熟禾提升效果最为显著。
3) 土壤团聚体稳定性受土壤性质与植被因子综合影响,其中MWD和GMD与孔隙度、有机碳、植被覆盖度呈极显著正相关。>2 mm团聚体比例、地上生物量和孔隙度是影响团聚体稳定性的主要因子,其对差异的解释率分别为56.10%、17.20%和2.50%。