


 下载:
下载:
-
毛叶木姜子(Litseamollis Hemsl.)是樟科(Lauraceae)木姜子属(Litsea)的多年生雌雄异株的落叶灌木或小乔木[1], 花期为3-4月, 果期为9-10月.枝叶和果实含挥发油, 具有明显的杀菌效果, 是我国重要的经济木本油料兼药用树种, 花黄色, 每一花序有花4~6朵, 先花后叶或与叶同时开放[1], 具有重要的观赏价值.其地理分布范围较广, 多生长在向阳山坡灌丛中或阔叶林中[1], 属于浅根性乡土树种, 耐瘠薄, 特别适应山地种植, 以野生资源为主[2], 在水土保持修复工程项目中, 作为主要生态恢复树种[3].
目前的研究主要集中在挥发油、表型特征、生物碱、药用贡献和扦插育苗[4-9]等方面, 由于对毛叶木姜子的生境情况和种群结构研究甚少, 对毛叶木姜子天然林的保护认识不足, 致使毛叶木姜子分布区越来越小, 群落退化严重, 种群数量急剧降低[10].目前市场上毛叶木姜子的原料主要来源于野生资源, 而野生资源的调查还处于起步阶段, 还需要进一步地完善毛叶木姜子的种群结构及种质资源资料, 才能进行种植资源的选优和保存[11].因此调查和研究毛叶木姜子种群结构, 探究开花情况及种群的年龄动态, 对于毛叶木姜子的种植和生产有重要作用.本研究以重庆市7个样地的毛叶木姜子天然林为对象, 从群落植物组成、种群结构、开花情况、种群生命表及存活曲线等方面进行研究, 旨在了解毛叶木姜子的生境情况及种群结构特征, 以期为重庆地区毛叶木姜子的种植生产和保护经营提供理论依据.
全文HTML
-
重庆市位于中国西南部、长江上游地区, 地处盆地东部, 地处105°11′-110°11′E, 28°10′-32°13′N之间的青藏高原与长江中下游平原的过渡地带, 地形由南北向长江河谷倾斜, 西北部和中部以丘陵、低山为主, 东南部靠大巴山和武陵山.属亚热带季风性湿润气候, 年平均气温16 ℃~18 ℃, 年平均相对湿度多在70%~80%, 在中国属高湿区, 年日照时数1 000~1 400 h, 日照百分率仅为25%~35%, 为中国年日照最少的地区之一.年平均降水量较丰富, 大部分地区在1 000~1 350 mm, 降水多集中在5-9月, 占全年总降水量的70%左右[12].调查涉及7个典型样地, 分布在巴南区、南川区、江津区和石柱土家族自治县4个区县, 涵盖了重庆市毛叶木姜子天然林分布的主要区域.各样地基本情况见表 1.
-
根据实际调查情况发现, 重庆市毛叶木姜子的盛花期在3月, 3月末开始花落, 因此在2018年3月选择保存较完整、集中连片分布的毛叶木姜子天然林开展调查, 能够保证个体已经开花且能较好地记录开花特征和性比.以30 m×30 m为乔木样方面积, 在每个样地内随机选择5个5 m×5 m的灌木样方, 再在每个灌木样方中设置2个1 m×1 m草本样方, 每个调查样地内共计设置35个灌木样方和70个草本样方[13].内容包括: ①每个样地的地点、坡度、坡向、海拔、经纬度、土壤类型、土壤厚度、干扰程度、群落郁闭度和植物名称等; ②采用每木调查法, 调查样方内所有开花的毛叶木姜子, 记录胸径、高度、冠幅、二级花枝数量、花枝长度和花序数量等, 没有胸径的植株需要记录除胸径以外的所有指标[14-15].实际调查典型样地中共计223株毛叶木姜子.
-
采用立木级结构代替年龄结构.根据毛叶木姜子的习性, 以胸径间隔(dBH)2, 4, 6 cm……为径阶距, 将个体划分为7个径级: Ⅰ级(dBH≤1 cm)、Ⅱ级(1 cm<dBH≤3 cm)、Ⅲ级(3 cm<dBH≤7 cm)、Ⅳ级(7 cm<dBH≤13 cm)、Ⅴ级(13 cm<dBH≤21 cm)、Ⅵ级(21 cm<dBH<31 cm)、Ⅶ级(dBH≥31 cm)[17].高度级(h)划分为7级: 1级(h≤2 m)、2级(2 m<h≤4 m)、3级(4 m<h≤6 m)、4级(6 m<h≤8 m)、5级(8 m<h≤10 m)、6级(10 m<h≤12 m)、7级(h>12 m).冠幅级(C)划分为7级: 1级(C≤2.5 m2)、2级(2.5 m2<C≤5 m2)、3级(5 m2<C≤10 m2)、4级(10 m2<C≤20 m2)、5级(20 m2<C≤30 m2)、6级(30 m2<C≤40 m2)、7级(C>40 m2). 3种分级互相独立[18-19].
-
静态生命表又称特定时间生命表, 根据在某一特定时间调查获得的种群各龄级的个体数编制, 能够提供种群各龄级数量变化的一般轮廓, 适用于寿命较长的木本植物[20].统计毛叶木姜子的个体数, 按照静态生命表的编制方法, 计算各个参数.其中: x为按年龄的分段径级; nx为x径级开始时的存活数; nx*为nx修正值; lx为x径级开始时的标准存活数, lx=nx/n0×1 000; dx为从x径级到x+1的径级死亡数, dx=nx-nx+1; qx为x径级的死亡率, qx=dx/nx; lx为从x径级到x+1径级的平均存活数, lx=(nx+nx+1)/2; Tx为x径级及以上各径级的存活个体总数, Tx=∑lx; ex为进入x径级个体的生命期望, 即平均能活的年限, ex=Tx/nx[17-19].为避免生命表中死亡率出现负值的情况, 采用匀滑技术对各龄级内的实际个体数进行匀滑修正[17, 21].
-
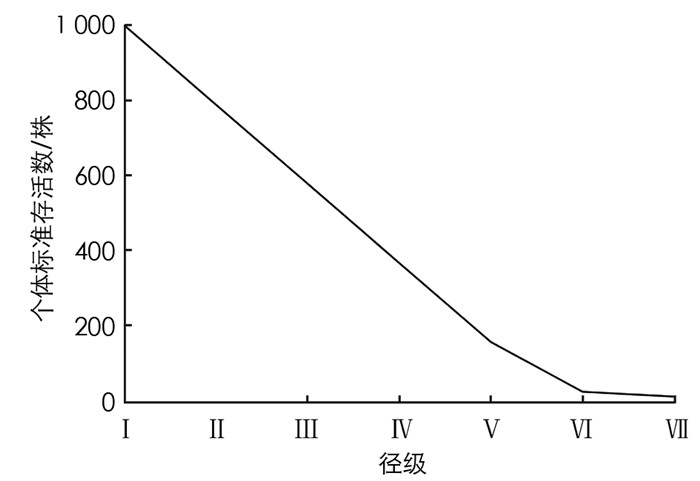
存活曲线能够直观地反映出存活率和死亡率随年龄变化的情况[20].根据Deevey对存活曲线的划分: Ⅰ型为凸型, 表示在接近生理寿命前只有少数个体死亡; Ⅱ型为对角线型, 各年龄阶段死亡率相等; Ⅲ型为凹型, 幼年期死亡率很高[20].以个体标准存活数为纵坐标, 以龄级为横坐标绘制毛叶木姜子种群存活曲线[21].
-
本研究针对毛叶木姜子天然林群落进行调查研究, 记录植物种的名称根据《中国植物志》[1]确定, 土壤类型参照《中国土壤分类系统表》[13].采用Microsoft Excel 2010和IBM SPSS Statistics 21进行数据统计分析和作图.
2.1. 样地设置及调查
2.2. 资料整理和数据分析
2.2.1. 种群年龄结构划分
2.2.2. 种群静态生命表的编制
2.2.3. 存活曲线的绘制
2.2.4. 数据处理
-
在调查的7个样地中, 共记录到维管束植物111种(不包括毛叶木姜子), 隶属于54科92属, 其中的包括蕨类植物7科8属8种, 裸子植物3科4属4种, 被子植物44科80属99种(表 2).各样地中以木本植物为主, 包括木本植物40科68属87种, 占总种数的78.38%, 草本植物14科24属24种, 占总种数的21.62%[10].含种数最多的科为蔷薇科(Rosaceae), 按种数由多到少的顺序为:山茶科(Theaceae)、荨麻科(Urticaceae)、樟科(Lauraceae)、菊科(Compositae)、禾本科(Gramineae), 含种数最多的属为悬钩子属(Rubus), 按种数由多到少的顺序为:栎属(Quercus)、柃木属(Eurya)、樟属(Cinnamomum)、山黄麻属(Trema)、山茶属(Camellia)、山矾属(Symplocos)和榕属(Ficus).
样地中出现次数较多的植物种类有马尾松(Pinusmassoniana Lamb.)、山莓(Rubuscorchorifolius L. f.)、芒(MiscanthussinensisAnderss.)、芒萁(Dicranopterisdichotoma (Thunb.) Berhn.)、毛竹(Phyllostachysheterocycla (Carr.) Mitford ‘Pubescens’)、杉木(Cunninghamialanceolata (Lamb.) Hook.)、狗脊(Woodwardia japonica (L.f.) Sm.)和柃木(Eurya japonicaThunb.), 其中马尾松在调查的6个样地中均有分布, 山莓、芒、芒萁在5个样地中有分布, 毛竹、杉木和狗脊出现在4个样地中.
-
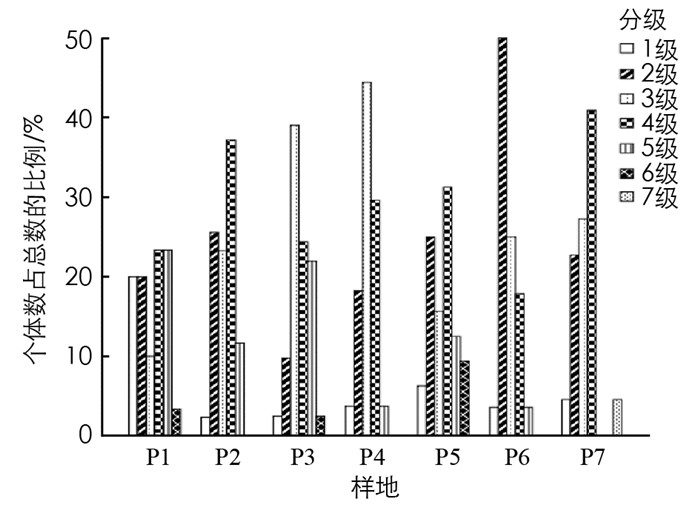
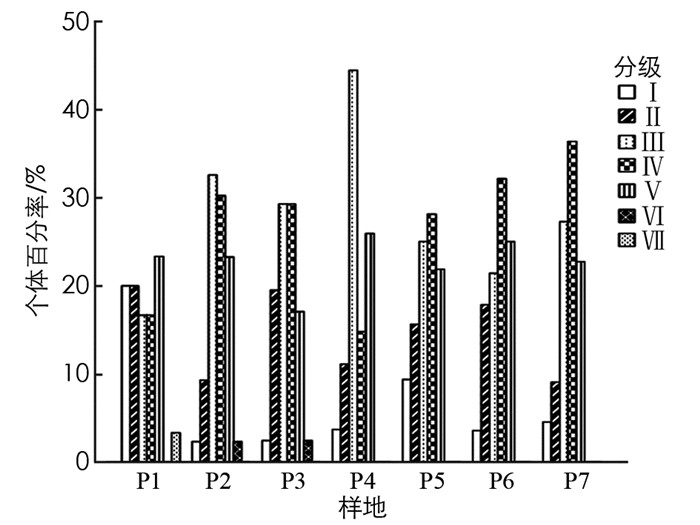
植物种群高度可以描述种群的垂直结构, 表现个体在群落垂直结构中的作用, 对高度结构的研究可以说明种群特点及演替趋势[22].各样地毛叶木姜子种群的高度结构见图 1, P1中高度主要分布在6~10 m之间; P2中高度主要在6~8 m之间, 第4级个体数最多, 占总数的37.21%; P3中高度主要在4~6 m之间, 个体较多的是第3级, 占总数的39.02%, 第1和第6级的个体数分布最少; P4中高度主要在4~6 m之间, 个体数较多的也是第3级, 占总数的44.44%; P5中包含1-6级, 个体分布相对均匀, 其中个体数最多的是第4级, 占总数的31.25%; P6中高度集中在2~4 m之间, 个体数最多的是第2级, 占总数的50.00%; P7中第4级分布个体数较多.对比各样地的高度级分布发现, 差异较为明显, 6个样地缺少第7级个体, 4个样地缺少第6级个体, P7缺少了第5和第6级个体, 且样地中第2, 3, 4级个体数量较多.
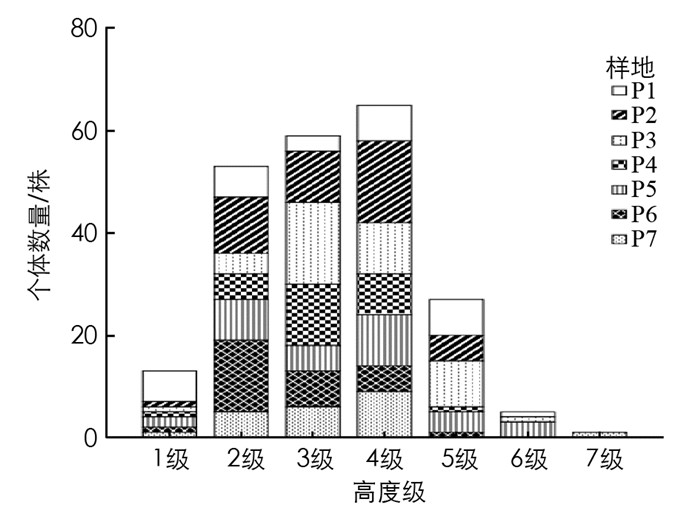
由图 2可知, 毛叶木姜子种群高度结构比较完整, 但是每个高度级之间差异较大, 主要集中在2-4级中, 共有177株, 占调查总数79.37%, 其中处于第4级的个体较多, 共有65株, 占总数的29.15%, 处于5-7级的有33株, 占总数的14.80%, 处于第1级的个体有13株, 占总数的5.83%.表明高度级位于中间的植株数量相对较多, 高度主要集中在2~10 m, 高度级大或小的个体数量较少.
-
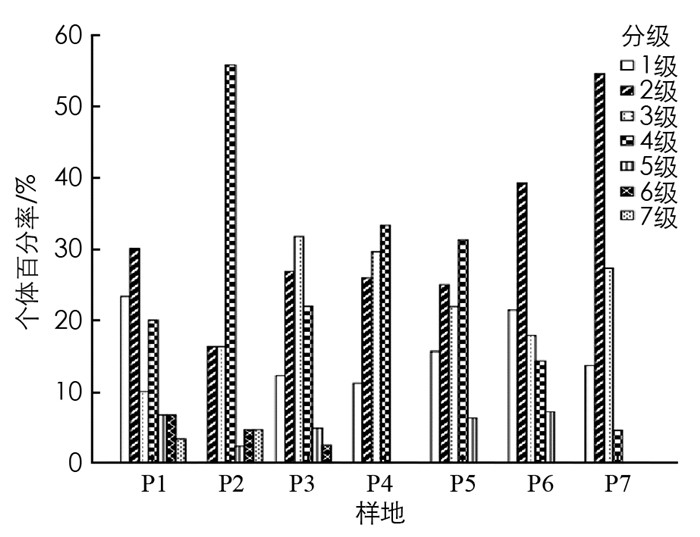
植物种群冠幅在一定程度上表明了树木的同化能力及空间占据和利用能力[19].各样地毛叶木姜子种群的冠幅特征见图 3, P1中冠幅级分布完整主要在2.5~5 m2之间, 第1和第2级个体数占总数的53.33%; P2中冠幅主要在10~20 m2之间, 占总数的55.81%; P3中个体主要分布在第2和第3级, 占总数的58.54%; P4中冠幅主要在2.5~20 m2之间; P5中个体数最多的是第4级, 占总数的31.25%; P6中冠幅主要在2.5~5 m2之间, 个体数最多的是第2级, 占总数的39.29%; P7中缺少第5, 6, 7级, 且第2级分布个体数较多.对比各样地的冠幅特征, 5个样地缺少第7级个体, 4个样地缺少第6级个体, 且样地中第2, 3, 4级个体数量较多.
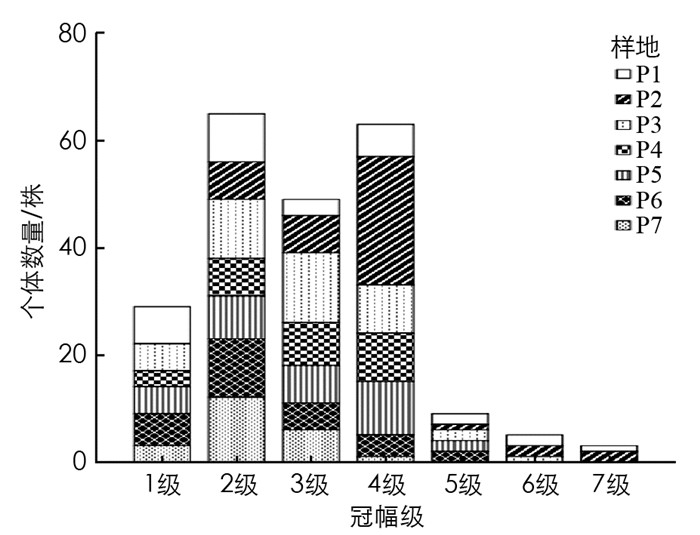
由图 4可知, 个体数最多的是第2冠幅级, 共有65株, 占总数的29.15%, 冠幅主要集中在2.5~20 m2, 每个冠幅级之间差异较大.第1和第2级共有94株, 占总数的42.25%, 第3和第4级共有112株, 占总数的50.22%, 在第5-8级中个体数量较少, 共有17株, 占总数的7.62%.将第1和第2级看作幼苗、幼树, 第3和第4级看作小树、5-7级看作成年树, 可见冠幅随级数升高个体数大致呈减少趋势.
-
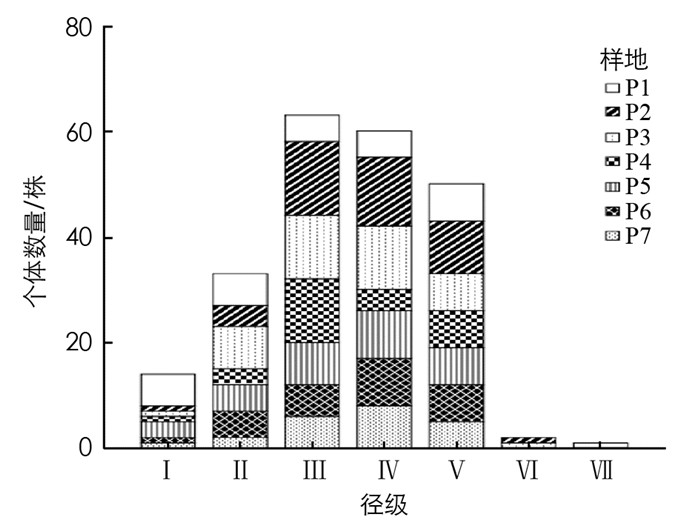
种群的径级结构可以代替年龄结构反映种群的生存状态[19].根据对胸径级的划分, 得到毛叶木姜子的径级结构图.各样地毛叶木姜子种群的径级结构见图 5, 每个样地中的径级结构差异较大, P1中胸径处于13~21 cm的个体最多, 占样地总数的23.33%; P2中胸径主要在3~13 cm之间, 在Ⅲ级的个体数最多, 占样地总数的32.56%; P3中胸径主要在3~13 cm之间, 占样地总数的58.54%; P4中胸径主要在3~7 cm之间; P5中个体数最多的是第Ⅳ级, 占样地总数的28.13%; P6中个体数最多的是第Ⅳ级, 占样地总数的32.14%; P7中缺少第Ⅵ和Ⅶ级, 且第Ⅳ级分布个体数较多.对比各样地的径级结构, 6个样地缺少第Ⅶ级个体, 5个样地缺少第Ⅵ级, 且各样地中第Ⅲ和第Ⅳ级个体数量较多.
由图 6可知, 毛叶木姜子种群的径级结构整体表现为中部较宽, 基部较窄, 顶部缺失的形式, 其中处于第Ⅰ级的个体数有14株, 占总数的6.28%, 第Ⅱ级的个体数有33株, 占总数的14.80%, 第Ⅲ级的个体数有63株, 占总数的28.25%, 第Ⅳ级的个体数有60株, 占总数的26.90%, 第Ⅴ级的个体数有50株, 占总数的22.42%, 第Ⅵ和第Ⅶ级总个体数为3株, 占总数的1.35%.种群胸径范围主要集中在3~13 cm, 幼龄和老龄个体较少, 以中龄个体为主, 种群结构较不稳定, 呈衰退型.
-
开花物候可影响植物的生殖成功, 在许多对于开花与结果的关系研究中表示花枝数越多, 开花数和座果数越多[23].根据对毛叶木姜子的枝数量、花序密度及性比调查, 各样地毛叶木姜子开花情况见表 3. P5和P3的个体平均二级花枝数量超过1 000, 平均花序密度大于2朵/cm, 开花情况好. P1和P2的个体平均二级花枝数量超过700, 平均花序密度在1.320~1.475朵/cm, 开花情况较好. P6和P7的个体平均二级花枝数量在374~262之间, 平均花序密度小于1.00朵/cm, 开花情况较差.对应各样地的性比, 雌雄个体数量相近的样地具有较好的开花情况, 雌雄个体数量相差较大的样地开花情况较差.
-
种群静态生命表能够反映种群的数量动态变化趋势及结构特征, 生命期望值(ex)反映的是x龄级开始时林木个体的平均期望生存能力[21-23].由表 4可知, 毛叶木姜子种群不同径级个体间的存活数差异较大, 总体为表现为中部较宽、顶部缺失.经匀滑技术对各龄级内的实际个体进行匀滑修正后, 得到毛叶木姜子种群静态生命表.在第Ⅵ级的死亡率为最高, 其次是第Ⅴ级, 死亡率最小的是第Ⅰ级, 随着龄级的增长, 死亡率变化波动较大, 呈现出随径级增大逐渐增加的趋势, 环境筛选及人类活动对其有较大的影响.种群的生命期望值最高的是第Ⅰ级, 其次是第Ⅱ和第Ⅲ级, 反映这3个阶段的生存能力较强, 生长状况较好, 生理活动相对旺盛, 第Ⅴ级生命期望值最低.由于受到人为活动的影响, 该种群第Ⅰ和第Ⅱ级的正常生长受到较大的限制, 加上种间竞争作用使得第Ⅳ和第Ⅴ级的植株被淘汰.
-
据图 7所示, 曲线为凹型和直线型之间, 不能判定是Deevey-Ⅱ型还是Deevey-Ⅲ型.在检验Deevey-Ⅱ型和Deevey-Ⅲ型时, 采用指数方程nx=n0e-bx来描述Deevey-Ⅱ型存活曲线, 用幂函数nx=n0x-b描述Deevey-Ⅲ型存活曲线[19, 24-25], 不同种群存活曲线的拟合方程: y=3 902.7e-0.755x(R2=0.905 5, F=47.941, P=0.001) y=2 505.3x-2.117(R2=0.714 7, F=12.528, P=0.017)
通过拟合对比所得方程可知, 指数方程模型的R2和F值均为最大, 由此看来, 毛叶木姜子种群存活曲线更符合Deevey-Ⅱ型.
3.1. 群落的植物种类组成
3.2. 种群结构分析
3.2.1. 种群高度结构
3.2.2. 种群冠幅特征
3.2.3. 种群径级结构
3.2.4. 开花情况及性比分析
3.3. 种群静态生命表及存活曲线分析
3.3.1. 种群静态生命表分析
3.3.2. 存活曲线分析
-
在国内对于花椒、茶树等经济林木, 关于产量、种植、种质资源等方面已经有了较为深入的研究[26-27].毛叶木姜子作为经济植物, 逐渐形成产业化, 但在过程中缺少对种群结构、开花情况及果实出油率的研究.根据对重庆市7个典型样地的毛叶木姜子的调查, 分析探讨毛叶木姜子的种群结构特征和开花情况等.结果表明, 群落中包含54科92属111种维管束植物, 主要为被子植物, 占总种数的89.19%.各样地中以木本植物为主, 共包括木本植物40科68属87种, 占总种数的78.38%, 可见毛叶木姜子群落的物种组成比较丰富.根据对毛叶木姜子种群结构的分析, 大多个体处于第2-4高度级, 占调查总数75.34%;大多数个体处于第2-4冠幅级, 占总数的79.37%;大多个体处于第Ⅲ-Ⅳ径级, 占总数的55.16%. P3和P5中个体高度集中在4~6 m, 冠幅较大, 雌雄数量基本相等, 开花情况最好.对应各样地的性比, 雌雄个体数量相近的样地具有较好的开花情况, 雌雄个体数量相差较大的样地开花情况较差.经过对种群静态生命表及存活曲线分析, 可知该种群属于衰退型种群, 第Ⅴ龄级生命期望值最低, 第Ⅵ龄级具有最高的死亡率, 推测受到人为活动的影响使该种群的正常生长受到较大的限制.经过拟合对比, 毛叶木姜子的存活曲线更符合Deevey-Ⅱ型.
在调查中发现, 毛叶木姜子的生长现状并不乐观, 人为干扰是引起种群结构和小生境改变的主要原因, 也在一定程度上改变了表型特征[2].早期的单一造林模式和不完善的经营管理, 造成毛叶木姜子人工林经济价值下降、遗传多样性降低[2].对于毛叶木姜子生境情况和种群结构的研究, 旨在能够提高对毛叶木姜子种群的认识, 有利于毛叶木姜子天然林的合理利用与保护, 并选择优质的种质资源进行保存, 为今后毛叶木姜子的种植生产和保护提供理论依据和参考.毛叶木姜子是多年生雌雄异株的落叶植株, 对开花特征和性比分析, 为结实率及果实出油量的研究提供理论基础.本次调查中由于受实验条件、时间等客观因素的制约, 仅对毛叶木姜子的种群结构及开花情况进行了研究, 而关于其花期时长、结实率和果实出油率等方面内容未涉及, 这也将成为未来进行深入研究的方向.为了使毛叶木姜子优质的野生资源得以保存, 建议在毛叶木姜子集中连片生长的区域, 尽量减少人为砍伐活动, 开展对于毛叶木姜子选种、扩繁和栽培技术的相关研究, 推广适用的应用技术.