

 下载:
下载:
-
张量的特征值和特征向量是矩阵的特征值和特征向量的推广,在自动控制、数据统计、优化、图像处理、固体力学、量子纠缠和高维马尔科夫链中都有重要的应用[1-9].复数域上的m阶n维的张量定义为
矩阵伪谱是分析矩阵特征值扰动的重要工具,在控制理论、微分方程数值解等领域都有着举足轻重的应用[10-11].张量伪谱可以看成是矩阵伪谱的推广,它与齐次动力系统的稳定解息息相关[12].
令zm-1=λ,若Re(z) < 0,则齐次动力系统
存在渐近稳定解.为了进一步判断其动力系统是否存在渐近稳定解,利用张量伪谱的定义,文献[12]得到了张量伪谱的圆盘定理,张量伪谱圆盘定理中得到的区域包含所有张量伪谱中的特征值,并且经常被用来判断齐次动力系统渐近稳定解的存在性.
本文通过对张量伪谱的进一步研究,得到了包含张量伪谱的更小的新包含域,新包含域包含所有张量伪谱中的特征值.数值例子验证了结果的有效性.
令
$ {r_i}(\mathit{\boldsymbol{A}})= \sum\limits_{\left( {{i_2}, \cdots , {i_m}} \right) \ne (i, \ldots , i)} {\left| {{a_{i{i_2}}} \cdots {i_m}} \right|} , r_i^j(\mathit{\boldsymbol{A}})= \sum\limits_{\left( {{i_2}, \cdots , {i_m}} \right) \ne (i, \cdots , i)} {\left| {{a_{i{i_2} \cdots {i_m}}}} \right|} - \left| {{a_{ij \cdots j}}} \right|$ ,可得如下张量伪谱的新包含域:定理1 设A=(ai1…im),E=(ei1…im)∈C[m,n],ε≥0.则
其中
证 令λ∈Λε(A),设非零向量x∈
$\mathbb{C} $ n是特征值λ对应的特征向量,即令|xp|=max{|xi|:i∈N},则|xp|≠0,其中N={1,2,…,n}.
若向量x中只有xp≠0,由(3)式可得
在等式(4)两边同时取绝对值,有
由Hölder不等式,有
由(5)式可得
即λ∈Λε(A)⊆Δ1(A)⊆Δ(A).
若向量x中至少有两个元xp≠0,xq≠0(p≠q),则有
求解xpm-1,有
在等式(6)两边同时取绝对值,有
即
即λ∈Λε(A)⊆Δ2(A)⊆Δ(A).
定理2 设A=(ai1…im),E=(ei1…im)∈C[m,n],ε≥0,则Δ(A)⊆D(A).
证 情形1 若λ∈Λε(A)⊆Δ1(A),由定理1可得
即λ∈D(A).
情形2 若λ∈Λε(A)⊆Δ1(A),由定理1知,存在p≠q,使得
又因为
则
即
若
$\left|a_{{p q} \cdots {q}}\right|\left(r_{p}(\boldsymbol{A})+n^{\frac{m-1}{2}} \varepsilon\right)=0 $ ,则有即有λ∈D(A).若
$ \left|a_{p q \cdots q}\right|\left(r_{p}(\boldsymbol{A})+n^{\frac{m-1}{2}} \varepsilon\right)>0$ ,则有即
或者
也就是说,λ∈Dp(A)或者λ∈Dq(A).即λ∈D(A).
注1 由定理2的结论可知,定理1得到的张量伪谱新包含域比文献[12]中定理3.1的包含域好,但是表达式更复杂,计算量更大.
我们用数值例子来说明结果的有效性.
其余的aijkl=0.令zm-1=λ,ε=0.4,0.5,0.8,1,在图 1中,我们给出关于z的包含区域,文献[12]的定理3.1中张量伪谱区域D(A)用黑色表示,定理1中张量伪谱区域Δ(A)用红色表示.由图 1可以看出,定理1的结果比文献[12]中定理3.1的结果好.
Pseudo-spectrum Localization Sets of Tensors
-
摘要: 张量伪谱可以看成是矩阵伪谱的推广,它在齐次动力系统中有着重要的作用.对张量伪谱圆盘定理进一步研究.利用张量伪谱中特征向量的最大元,给出了张量伪谱的新包含域.数值例子验证了结果的有效性.Abstract: The definition of pseudo-spectrum of tensors can be seemed as a generation of the definition of pseudo-spectrum of matrices, which plays important role in the homogeneous dynamical system. The pseudospectra localization of tensors is often used to determine the existence of its asymptotically stable solution for the homogeneous dynamical system. In this paper, a new pseudospectra localization of tensors is established by the maximum element of the eigenvector. Numerical experiments are reported to show effectiveness of our results.
-
Key words:
- tensor /
- pseudo-spectrum /
- localization set .
-

-
[1] QI L Q.Eigenvalues of a Real Supersymmetric Tensor[J].Journal of Symbolic Computation, 2005, 40(6):1302-1324. doi: 10.1016/j.jsc.2005.05.007 [2] doi: http://d.old.wanfangdata.com.cn/Conference/8576630 LI C Q, LI Y T, KONG X.New Eigenvalue Inclusion Sets for Tensors[J].Numerical Linear Algebra with Applications, 2014, 21(1):39-50. [3] LI C Q, CHEN Z, LI Y T. A New Eigenvalue Inclusion Set for Tensors and Its Applications[J].Linear Algebra and its Applications, 2015, 481:36-53. doi: 10.1016/j.laa.2015.04.023 [4] LI C Q, WANG F, ZHAO J X, et al.Criterions for the Positive Definiteness of Real Supersymmetric Tensors[J].Journal of Computational and Applied Mathematics, 2014, 255:1-14. doi: 10.1016/j.cam.2013.04.022 [5] BU C J, WEI Y P, SUN L Z, et al.Brualdi-Type Eigenvalue Inclusion Sets of Tensors[J].Linear Algebra and its Applications, 2015, 480:168-175. doi: 10.1016/j.laa.2015.04.034 [6] BU C J, JIN X Q, LI H F, et al.Brauer-Type Eigenvalue Inclusion Sets and the Spectral Radius of Tensors[J].Linear Algebra and its Applications, 2017, 512:234-248. doi: 10.1016/j.laa.2016.09.041 [7] 桑彩丽, 赵建兴.非负矩形张量最大奇异值的S-型上界[J].西南师范大学学报(自然科学版), 2018, 43(6):1-5. doi: http://d.old.wanfangdata.com.cn/Periodical/xnsfdxxb201806001 [8] 钟琴.非负矩阵最大特征值的新界值[J].西南大学学报(自然科学版), 2018, 40(2):40-43. doi: http://d.old.wanfangdata.com.cn/Periodical/szjsyjsjyy200204007 [9] 钟琴, 周鑫, 牟谷芳.非负矩阵谱半径的上界估计[J].西南大学学报(自然科学版), 2017, 39(6):50-53. doi: http://d.old.wanfangdata.com.cn/Periodical/xnsfdxxb201602002 [10] TREFETHEN L N. Spectra and Pseudospectra:The Behavior of Nonnormal Matrices and Operators[J].Bulletin of the American Math Society, 2005(2):277-284. [11] BOSE N. Test for Lyapunov Stability by Rational Operations[J]. IEEE Trans Automat Control, 1975, 20(5):700-702. doi: 10.1109/TAC.1975.1101045 [12] CHE M L, LI G Y, QI L Q, et al.Pseudo-Spectra Theory of Tensors and Tensor Polynomial Eigenvalue Problems[J].Linear Algebra and its Applications, 2017, 533:536-572. doi: 10.1016/j.laa.2017.07.026 -
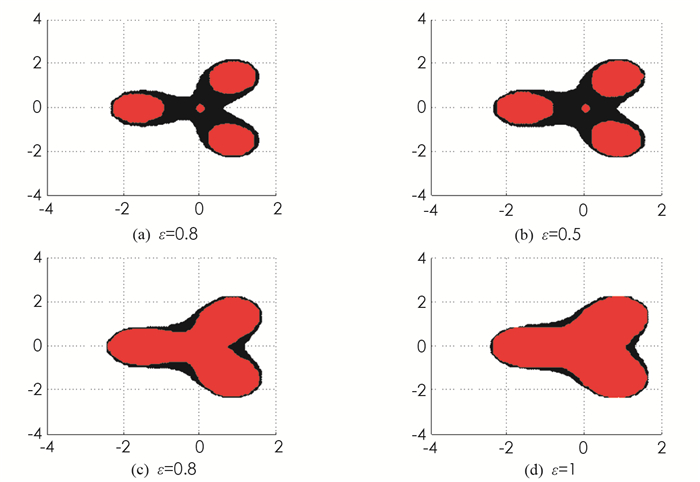
图( 1)
计量
- 文章访问数: 1894
- HTML全文浏览数: 1663
- PDF下载数: 215
- 施引文献: 0