


 下载:
下载:
-
反应扩散方程一直是生物种群数学模型研究中比较经典的模型之一,可以很好地解释生物种群在现实世界中存在的模式[1-10].但是近年来,由于在种群建模的过程中既不能忽视种群走动过程中的扩散效应,也不能忽视种群相互作用过程中的时滞因素,所以考虑非局部效应对种群分布模式的影响变得越来越重要.
全文HTML
-
英国的生物数学学家Britton[11]在logistic单种群模型的基础上提出存在非局部效应的单种群模型,主要考虑了种群在空间中扩散之后,种群密度的变化率可能受到过去某段时间内和相邻区域的种群相互作用的影响,即种群的非局部效应对种群时空分布的影响.该带有非局部效应的单种群模型[12]表示如下:
其中:(f**u)(x,t)=∫-∞t∫
$\mathbb{R}$ f(x-y,t-s)u(y,s)dyds;(x,t)∈$\mathbb{R}$ ×(0,+∞);参数d,r,α,β均是正参数,r表示种群的内禀增长率,α表示种群的聚集效应,β表示种群个体空间竞争产生的效应,d表示种群的扩散系数;$\mathit{\Delta } = \frac{{{\partial ^2}}}{{\partial {x^2}}}$ 是一维空间中的拉普拉斯算子,而且1+α-β>0.在方程(1)中,其核函数其中τ表示种群个体反应的平均时滞.该模型(1)的各项都有着相应的生物学上的解释:αu表示种群的聚集作用、-βu2表示种群对空间的竞争、积分项-(1+α-β)(f**u)表示种群内食物资源引发的竞争,且核函数满足如下性质:
(H1) f∈L1(
$\mathbb{R}$ ×(0,∞))并且tf∈L1($\mathbb{R}$ ×(0,∞)),其中f∈L1($\mathbb{R}$ ×(0,∞))表明卷积f**u是关于时间和空间的可积函数.(H2) f满足f**1=1,也就是说∫
$\mathbb{R}$ ∫0∞f(x,t)dtdx=1成立.这个条件表明原系统的均匀状态的解是不会改变的,仍对应着局部形式下的系统的均匀平衡态.(H3) 因为f为权函数,故核函数f(x,t)≥0.
(H4) 核函数f刻画着u(y,s)关于u(x,t)(s≤t)的从x到y的非局部效应.
文献[12]主要讨论了该单种群模型的3类分支现象和其所对应的周期驻波解、空间周期结构的解的存在性.
-
当选择
$f\left( {x, t} \right) = \frac{1}{{\sqrt {4\pi t} }}{{\rm{e}}^{ - \frac{{{{\left| x \right|}^2}}}{{4t}}}}\frac{1}{\tau }{{\rm{e}}^{ - \frac{t}{\tau }}}$ 时,在变换中令
有
依据文献[13],模型(1)被转化为如下的方程组
其中
本节主要通过分析系统(2)来得到系统(1)的动力学行为.系统(2)有3个平衡点(0,0),
$\left( { - \frac{1}{\beta }, - \frac{1}{\beta }} \right)$ ,(1,1).但是从生物种群的角度,主要关注系统(2)在第3个平衡点(1,1)附近的动力学行为,因为该平衡点对应着种群生存环境容纳量的平衡状态.系统(2)在平衡点(1,1)处线性化后得到如下的线性系统:其中a11=r(α-2β),a12=-r(1+α-β),a21=
$\frac{1}{\tau }$ ,a22=$-\frac{1}{\tau }$ .这里为了研究方便假设系统(2)满足如下的初始条件,
且有光滑的边界条件,进一步系统(2)边界条件为纽曼边界条件,即
其中n表示边界∂Ω的外法向量.
根据线性系统(3)和文献[7],令
并带入到方程(3)得到如下的方程式,
也就是
为了找到非零的常数α1和α2,可以得到如下的特征方程,
该特征方程等价于
其中
依据特征方程(4),得到两个特征根为:
综合上面的分析可得系统(2)的平衡点(1,1)渐进稳定的充分条件和在该平衡点处形成空间均匀周期解的条件.
定理1 当
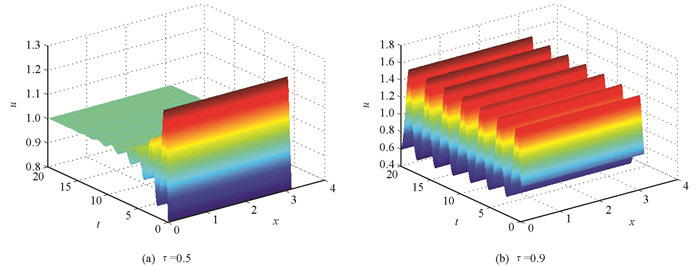
$\tau < \frac{1}{{r\left( {\alpha - 2\beta } \right)}}$ ,4d(a11a22-a12a21)>(a11+a22d)2时,特征方程(4)对于所有的k而言,其根的实部都是小于零的,则平衡点(1,1)是渐进稳定的;当$\tau = \frac{1}{{r\left( {\alpha - 2\beta } \right)}}$ ,4d(a11a22-a12a21)>(a11+a22d)2时,特征方程(4)在k=0时有一对纯虚根,并且对任意的k>0,其根的实部都是小于零的,则平衡点(1,1)将失稳而发生空间均匀的Hopf分支,形成空间均匀的周期解.依据定理1选取参数d=1.0,r=3.0,α=1.2,β=0.4,当τ=0.5时,系统(2)的平衡点(1,1)是渐进稳定的(图 1(a));当τ=0.9时,系统(2)的平衡点(1,1)将失去稳定性,在此处诱发空间均匀的周期解(图 1(b)).
从上面的数值计算结果来看,种群模型(2)在一定的条件下存在空间均匀的解,这种解要么为一个均匀的稳态,要么是一个均匀的周期解.但是这种解是否会受到模型中其他参数的影响进而转化为其他形式的时空模式,将在下节中给予讨论.
-
在第2节中,主要借助空间均匀的Hopf分支理论和Hurwtiz准则得到系统诱发空间均匀周期解的条件,然后借助数值计算的方法,选取合适的系统参数,来验证前面所得到的理论分析.但是在模型(2)中还存在其他形式的时空分布模式.由定理1知,当时滞量a增大到d=0.001时,系统(2)将在平衡点r=3.0处诱发超临界的Hopf分支并诱发空间均匀的周期解形成.该解将会在扩散的影响下,有可能导致系统(1)形成周期行波解.
为了得到系统(1)周期行波解的存在性,需要把系统(2)转化为行波方程.对于系统(2)而言,若有u(x,t)=ζ(z),v(x,t)=ξ(z),z=x+ct,则可以得到关于变量ζ,ξ的行波方程如下:
令ζ′=
${\bar \xi }$ ,ζ′=${\bar \xi }$ ,于是系统(6)化为如下形式:系统(7)有两个与时滞量τ无关的平衡点(0,0,0,0)和(1,0,1,0).于是,将系统(7)在平衡点(1,0,1,0)处线性化,可得到相应的特征方程为:
令λ=iω0(ω0>0)代入到方程(8)中,同时分离实部和虚部有
由方程(9)有如下结果
于是,由方程(10)可得到时滞τ的临界值为
其中
方程(8)两边关于时滞τ微分可得
于是当τ=τ0时,
若r(1+α-β)-1>0,则Reλτ′>0.
综合以上分析,得到如下结论:
定理2 若r(1+α-β)-1>0和α-2β>0,那么当时滞参数τ通过其临界值τ0时,行波系统通过Hopf分支形成了一个小振幅的周期解,也就是系统的周期行波解.
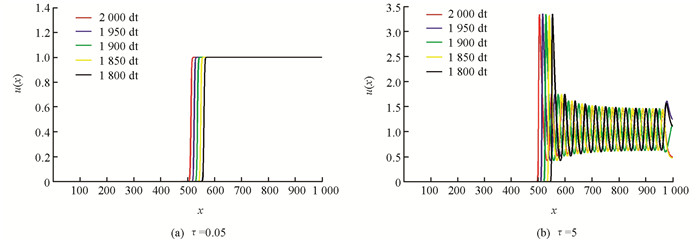
对系统(2)进行数值验证.选取一维的空间来反映该种群在空间的分布状况.在所有的数值结果中,系统的边界条件为Neumann边界条件且空间范围为0~1 000,时间方向上的范围为0~300,且空间步长Δx=1,时间步长Δt=0.05,并且初始条件为在系统平衡态上的扰动.取系统参数为d=1,r=3,α=0.5,β=0.2,系统在τ=0.05,τ=5时的数值结果见图 2.
从图 2的数值模拟结果可知当时滞量非常小的时候,系统存在单调递增的波前解.但是,随着时滞的增加,系统行波解的波形也随之发生变化,出现周期行波解.这些结果的生物解释和意义为:一定的情况下,现实生态环境中的种群在空间的分布会呈现出行波的传播方式.于是可以借助行波的波速和振幅,预测种群在空间传播的速度和传播的范围,为提前做好种群资源的管理,特别是某些有害入侵物种的控制提供一些理论方面的帮助和指导.
-
本文主要讨论了一类耦合了非局部效应的单种群模型的时空分布模式,主要包括空间均匀的周期解和空间非均匀的周期行波解的存在性,并借助数值计算的方法模拟出这些时空分布模式.所得到的结果主要从理论的角度说明了种群在空间分布的波动行为,虽然不能很好反映出现实世界中种群分布的现状,但是为生物学家分析某类入侵种群在空间分布上的传播速度和传播范围提供了一定的指导和借鉴.