



 下载:
下载:

-
草地贪夜蛾(Spodoptera frugiperda),又名秋黏虫、行军虫,起源于美洲,被联合国粮农组织称为“全球重大跨国迁飞性农业害虫”,对农作物的危害极大[1].其食谱广泛,主要包括禾本科、菊科和豆科植物,草地贪夜蛾幼虫喜食玉米、水稻和甘蔗等农作物,且主要啃食作物的幼嫩部分,具有严重的危害性.自2019年1月由东南亚入侵我国云南、广西以来,草地贪夜蛾随着温度的增加以及农作物的丰富逐渐向全国大面积迁飞,目前已迁飞至河南、山东等地区,对我国农作物产区存在着巨大的威胁. 2019年5月已在重庆的巫山、巫溪、云阳、北碚和潼南等地区发现有草地贪夜蛾出现并有迁飞的趋势,入侵到重庆的草地贪夜蛾对当地的玉米生产造成了严重影响(图 1).
草地贪夜蛾幼虫头部均具有“Y”字纹,尾部有4个呈正方形排列的黑点,幼虫期与其他亲缘关系较近的鳞翅目昆虫有较为明显的差别[2].野外采集的草地贪夜蛾主要分为“玉米型”(corn-strain)和“水稻型”(rice-strain)两个亚群体[3],这两种基因亚型无法从形态学上分辨,但它们在生理特征、行为特征和取食倾向上有着较大差别[4-5].此外,这两个基因型相互之间可以交配,产生杂合子[6].其亚型可根据目前最常用于鉴定草地贪夜蛾的分子条形码,基于线粒体细胞色素C氧化酶亚基I基因(cytochrome c oxidase subunit I,COI)和Z染色体上编码磷酸甘油醛异构酶基因(triose phosphate isomerase,Tpi)进行鉴定区分[7-12].本研究通过比较分析上述两个基因,鉴定了入侵重庆地区的草地贪夜蛾群体亚型,为开展防控以及分析草地贪夜蛾种群差异相关工作提供理论基础,对掌握侵入重庆市的草地贪夜蛾的遗传信息和制定相应的防治策略有重要意义.
全文HTML
-
本实验所用的草地贪夜蛾幼虫及成虫,由重庆市巫山县植保站、巫溪县植保站和潼南区植保站提供,共33只,其中巫山21只,巫溪2只,潼南10只.样品采集于2019年6月14日至2019年6月21日,其中死虫浸泡于75%酒精中,从巫山采集的活虫置于培养皿中,喂食新鲜玉米叶,带回实验室后迅速对其进行形态学观察,随后置于液氮速冻后,在-80 ℃温度下储存,用于后续研究.
-
用液氮将单头幼虫或成虫研磨成粉末状,以CTAB法提取DNA[13],DNA用NanoDrop ND-2000仪器测定浓度.于-20 ℃储存用于后续实验.
分别用COI和Tpi基因的引物[14]对所提取的草地贪夜蛾基因组DNA进行PCR扩增,基因组稀释到50 mg/L,使用Thermal scientific公司的Dream Taq酶进行扩增,扩增反应体系如下:Dream Taq预混液25 μL;上游引物/下游引物各2 μL;草地贪夜蛾基因组DNA 2 μL;补足ddH2O至50 μL. PCR反应体系为94 ℃,8 min;94 ℃,30 s;55 ℃,30 s;72 ℃,30 s,30个循环;72 ℃,10 min;用1.5%琼脂糖电泳检测,PCR产物进行测序分析.
-
在GenBank中下载已报道的草地贪夜蛾的COI和Tpi序列,使用MEGA 6.0进行多重序列比对,剔除冗余的低质量序列后,进行进化树分析.根据COI和Tpi基因具有“水稻型”和“玉米型”的亚型,用ClustalX(1.83)软件分析所得序列的差异位点,以判断重庆市分布的草地贪夜蛾的亚型.
1.1. 实验材料
1.2. DNA提取和PCR扩增
1.3. 测序结果分析
-
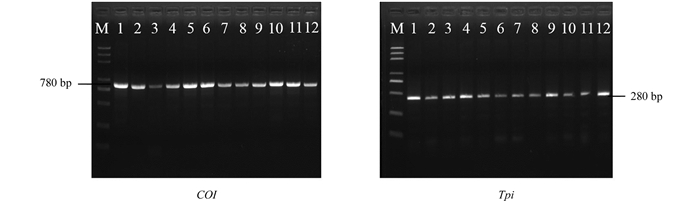
使用COI基因和Tpi基因的引物对提取的基因组进行PCR扩增,COI基因扩增产物约为780 bp,Tpi基因扩增产物约为280 bp,共22份样本可以扩增出目的条带(图 2),其中有17份样本两个基因均可扩增得到,部分样品只能扩增出其中一个基因,我们选择这17个样本的PCR产物进行测序分析.
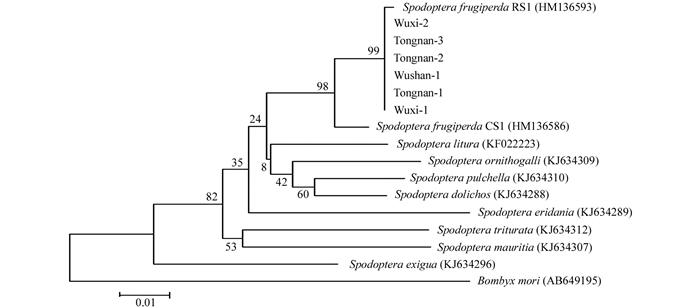
将得到的测序结果在NCBI中进行序列比对,下载与之相似度较高的序列,同时下载与草地贪夜蛾亲缘关系较近的鳞翅目昆虫,如斜纹夜蛾(Spodoptera litura)等的COI序列,以家蚕(Bombyx mori)的COI序列作为对照,用MEGA 6.0进行多重序列比对,并构建进化树.发现所有测序序列均与草地贪夜蛾聚为一枝,其中巫山组样品的测序结果完全一致,因此在后续进化树分析中以其中一组(Wushan-1)为代表(图 3).
-
将测序得到的COI基因序列与草地贪夜蛾“水稻型”COI基因(GenBank序列号:HM136593)进行比对发现,仅有2个样品与“水稻型”COI基因有一个碱基的差异,其余样品无差异.而所有样品与“玉米型”COI基因(GenBank序列号:HM136586)相比,有14个碱基的差异(图 4),对这些序列做进化树分析,发现序列全部归在了“水稻型”的基因型中(图 3),表明了这批样品的COI基因亚型是“水稻型”.
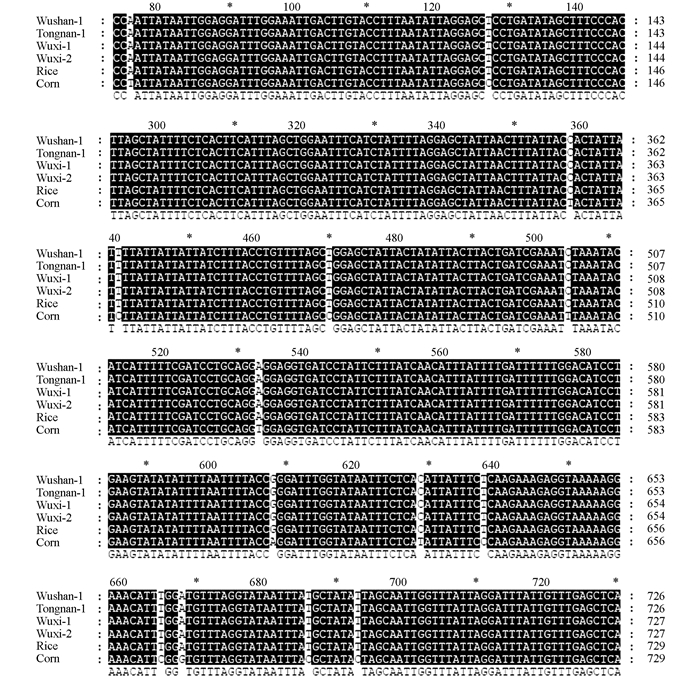
为了分析得到的Tpi基因序列具体的差异位点,用ClustalX (1.83)软件对序列进行比对分析,发现来自巫山和巫溪的样品中,124~125位的碱基既有GA也有AT (图 5),该位点是"水稻型"和"玉米型"的差异位点之一,表明入侵重庆的草地贪夜蛾可能存在"水稻型"与"玉米型"杂合型.
2.1. COI基因与Tpi基因的扩增
2.2. COI基因与Tpi基因亚型鉴定
-
已有的报道表明,我国大部分地区发现的草地贪夜蛾的Tpi基因亚型大多为“玉米型”,少量为杂合“水稻型”,而COI基因既有“玉米型”又有“水稻型”[15-16].在我们的研究中发现,入侵到重庆市巫山县、巫溪县和潼南区的草地贪夜蛾的COI基因仅存在“水稻型”亚型,说明迁飞到重庆的草地贪夜蛾的COI基因亚型多是“水稻型”. Tpi基因的亚型中,有相当一部分的“水稻型”亚型的存在(23.53%).此前在有关广州地区的报道中发现了杂合的“水稻型”Tpi基因,但比例很低,而在更早时期在云南有关草地贪夜蛾的鉴定没有发现“水稻型”Tpi基因[15-16],暗示着入侵重庆的部分草地贪夜蛾可能来源于广州地区,并在迁飞过程中“水稻型”亚型与“玉米型”亚型交配,产生了杂合的“水稻型”Tpi基因,但由于本研究样本量还不够大,还需要更大的样本量来确认重庆地区的Tpi基因“水稻型”亚型是否呈现较高比例.尽管如此,“水稻型”Tpi基因比例的升高值得引起我们的重视.
草地贪夜蛾的卵不滞育,无法越冬,需迁徙繁育.有研究表明,草地贪夜蛾可以在北纬29°到北纬30°的地区越冬[17],这暗示着草地贪夜蛾最北可以在四川省、重庆市、湖北省、湖南省、江西省、安徽省和浙江省的某些地区越冬.我国南方地区是水稻的盛产地,面对可能出现的“水稻型”亚型的草地贪夜蛾的比例升高,需要加强对水稻产区的监测.因此,如何在早期发现和鉴别草地贪夜蛾,并加强对草地贪夜蛾的防控是至关重要的.
目前,利用特定基因对物种进行分子鉴定是主流方法,COI和Tpi基因作为DNA条形码足以对草地贪夜蛾进行物种鉴定,可以快速鉴定当地是否有草地贪夜蛾的入侵,为快速应对草地贪夜蛾的入侵提供了基础.但是,对于草地贪夜蛾亚型的鉴定,仅仅依靠COI和Tpi这两个基因是不够的,由于这两个亚型之间不存在生殖隔离,它们交配会产生杂合型的基因,这对草地贪夜蛾亚型的判断会造成影响.因此,对于这两种亚型的鉴定还需要从多个角度来进行验证,比如分析不同亚型的肠道微生物组成,阐明不同亚型取食倾向差异的机制等.
本文初步探讨了入侵重庆的草地贪夜蛾的群体亚型,COI基因全为“水稻型”亚型,Tpi基因主要为“玉米型”亚型,存在杂合的“水稻型”亚型.因此,重庆市以及重庆以北省市未来对草地贪夜蛾的防控工作,不能仅仅局限于玉米产区,还要对水稻产区进行实时监控,长期监测草地贪夜蛾群体亚型的变化,警惕杂合“水稻型”亚型群体的扩大.