





-
开放科学(资源服务)标志码(OSID):

-
水分是植物正常生长的基础条件,但水分过多会打破植物体内的水平衡,影响正常生长,甚至导致死亡[1]. 我国长江流域冬油菜主产区秋季多阴雨,且实行稻油轮作,在油菜苗期土壤湿度大,易遭受湿害胁迫.
湿害是指土壤水分长期处于饱和状态而使植物受到危害. 湿害胁迫下,土壤含氧量不足,植物根系处于低氧或缺氧状态[2],活力下降[3],表面积、体积和平均直径增长受抑制[4];植株新叶形成受阻,叶片数量减少,蒸腾速率降低,叶绿素质量分数下降,光合作用的电子传递过程也受抑制,导致净光合速率降低[5-8]. 植物对湿害胁迫的响应主要表现在形态结构、生理生化和分子机制等方面. 其中,形态结构响应包括通气组织和不定根的形成、皮孔增生和叶柄偏上生长等[9-10];生理生化响应主要体现在糖酵解途径增强、活性氧自由基过多产生与清除以及低氧条件引发的能量危机[9, 11];分子机制响应包括基因表达调控和转录后修饰等方面,如好氧蛋白的合成受阻和厌氧蛋白的新合成等[12].
植物耐湿性是由多性状组成的复杂综合性状,不同基因型和生长发育的不同时期均有不同. 历年来,前人对油菜资源和品种进行了许多耐湿性鉴定. 尹继春等[13]采用定期灌水控制土壤湿度法鉴定了湿害对油菜不同生育时期的影响以及不同品种的耐湿性. 卢长明等[14]采用冷开水浸泡干燥的种子,测定发芽系数鉴定耐湿性,认为在缺氧条件下筛选发芽势强的基因型可能对改善耐湿性具有明显效果. 范其新[15]将露白种子淹水处理24 h后在滤纸上继续发芽,调查相对成苗率、活力指数、电导率和苗长等性状,根据活力指数对100份甘蓝型油菜种质的耐湿性进行了分类. 陈洁[16]将露白种子进行密闭淹水12 h后在沙床上继续发芽,6 d后测定相关性状,计算活力指数,鉴定了不同油菜种质的耐湿水平. 李真等[17]对甘蓝型油菜DH(Double Haploid)群体进行湿害处理(土壤表面保持2 cm水层),比较各性状的表现并计算耐湿系数,筛选出极端材料. 薛远超[18]采用萌发种子露白后淹水处理12 h再正常培养6 d,然后在苗期保持高于土面约3 cm水层的方法对EMS(Ethylmeth-anesulfonate)诱变的甘蓝型油菜种质进行了萌发期和苗期耐湿性鉴定. 陈娟妮等[19]在四叶一心时期进行人工淹水处理,鉴定了32个甘蓝型油菜品种的耐湿性. 张雪花等[20]将发芽种子密闭水淹处理14 h后继续培养6 d,进行油菜资源的耐湿性鉴定和筛选. 涂玉琴等[21]保持1~2 cm浅水层的湿害处理方式,对15份源自甘蓝型油菜与蔊菜杂交获得的创新种质进行了耐湿性鉴定和综合评价.
与其他作物相比,甘蓝型油菜种质资源的耐湿性综合评价较少,前人的研究多采用萌发期或苗期淹水法进行耐湿性鉴定,所用试验材料在数量上和来源上均具有一定的局限性. 本研究利用国内外广泛收集的200份甘蓝型油菜种质资源,采用盆栽方式进行苗期湿害胁迫(保持土壤最大持水量且表面无积水)和正常灌溉,测定形态生长和生理生化性状,并通过主成分分析、隶属函数分析和灰色关联度分析方法进行耐湿性综合评价,为耐湿性油菜新品种培育和油菜湿害机理研究提供参考.
HTML
-
将国内外多家油菜研究单位广泛收集的200份甘蓝型油菜资源材料,以盆栽方式种植于重庆市油菜工程技术研究中心歇马基地的旱棚内. 盆钵直径约25 cm,高约30 cm. 当油菜幼苗长至四叶一心时,选择长势均匀一致的盆钵进行湿害胁迫(Waterlogging,WL)和正常灌溉(Well watering,WW)处理. 采用浙江托普仪器有限公司的TZS-1K土壤水分测定仪定期测定土壤含水量,使WL组保持土壤最大持水量且表面无积水(土壤含水量约为30%),WW组土壤含水量保持在20%~23%的范围内.
-
处理4周后,进行性状调查. 幼苗心叶展平视为1片叶,绿色面积超过叶片总面积1/2视为绿叶. 调查WL组和WW组每个材料随机3株幼苗的绿叶数(Number of Green Leaves,NGL). WL组和WW组每个材料随机选取3株,称量地上部鲜质量(Shoot Fresh Weight,SFW),110 ℃杀青、75 ℃烘干至恒质量后称量地上部干质量(Shoot Dry Weight,SDW). WL组和WW组每个材料随机选择3株,取倒数第2叶、第3叶混合样品,采用硫代巴比妥酸法和硫酸-蒽酮比色法分别测定叶片丙二醛质量分数(Malonaldehyde Content,MDA)和可溶性糖质量分数(Soluble Sugar Content,SSC)[22],采用考马斯亮蓝-G250法测定叶片可溶性蛋白质量分数(Soluble Protein Content,SPC)[23],采用南京建成生物工程研究所研发的试剂盒A084-3和A001-1分别测定叶片过氧化物酶活性(Peroxidase Activity,POD)和超氧化物歧化酶活性(Superoxide Dismutase Activity,SOD).
-
描述统计和方差分析分别采用Microsoft Office Excel 2016和DPS V15.10[24]完成.
采用公式(1)计算NGL,SFW,SDW,SSC,SPC,POD和SOD的耐湿系数,公式(2)计算MDA的耐湿系数[25]. 采用公式(3)和公式(4)对各性状耐湿系数进行标准化,用欧式距离+类平均法进行系统聚类,用主成分分析法计算各主成分因子特征值、贡献率和累计贡献率,选取累计贡献率达80%以上的主成分因子数,按照公式(5)计算各主成分因子特征值,公式(6)计算主成分的综合值[24-25]. 利用公式(7)和公式(8)分别计算各性状隶属函数值和各材料的平均隶属函数值[25-26]. 根据李阳阳等[25]的方法选择各性状耐湿系数的最大值,构成理想型耐湿材料的耐湿系数,利用公式(9)和公式(10)分别计算各材料与理想型耐湿材料的关联系数和综合关联度.
上述公式中,n为试验材料数,m为测定性状数,p为所选取的主成分因子数;αij为第i个材料第j个性状的耐湿系数,tij为第i个材料第j个性状在WL组的性状测定值,Tij为第i个材料第j个性状在WW组的性状测定值;Sj为第j个性状耐湿系数的标准差,αj为第j个性状耐湿系数的平均数,αij′为第i个材料第j个性状的耐湿系数标准化值;Pik为第i个材料第k个主成分因子值,λkj为第j个性状第k个主成分因子的特征向量值,Pi为第i个材料的主成分因子综合值;Fij为第i个材料第j个性状耐湿系数的隶属函数值,αminj为第j个性状的最小耐湿系数值,αmaxj为第j个性状的最大耐湿系数值,Fi为第i个材料的平均隶属函数值;εij为第i个材料第j个性状与理想型耐湿材料的关联系数,Ij为理想型耐湿材料第j个性状的耐湿系数值,
$\mathop {\min }\limits_i \mathop {\min}\limits_j \left| {{I_j} - {\alpha _{ij}}} \right|$ 为二级最小差,$\max _i \max _j\left|I_j-\alpha_{i j}\right|$ 为二级最大差,ρ为分辨系数(在此取值0.5),Ci为第i个材料的综合关联度.
1.1. 试验材料与种植
1.2. 性状测定
1.3. 数据分析
-
由表 1和表 2可知,与正常灌溉相比,湿害胁迫使甘蓝型油菜NGL,SFW,SDW,SPC和SOD极显著降低,MDA,SSC和POD极显著升高(p<1%).
-
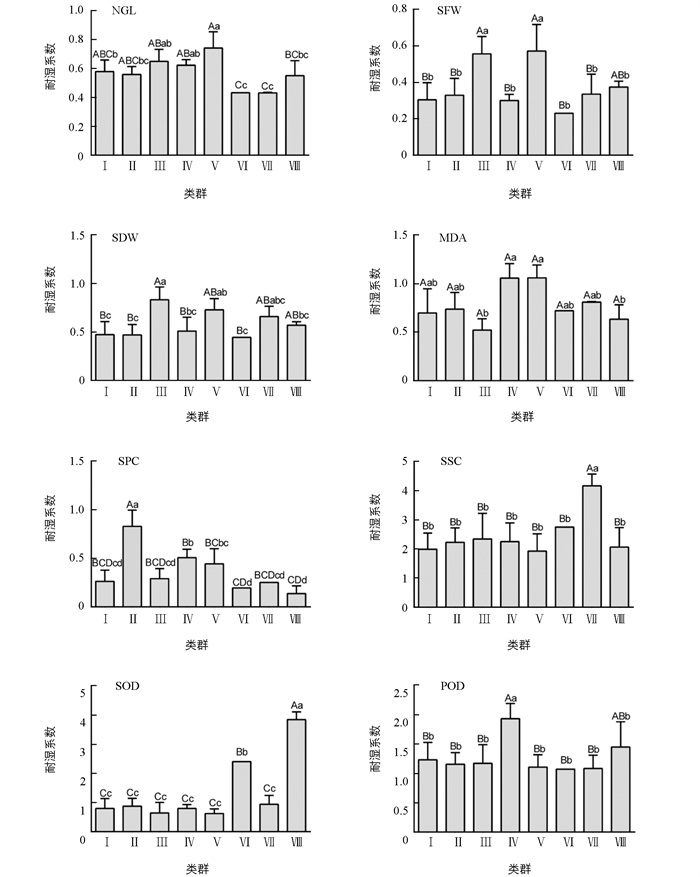
通过聚类分析,将200份材料分为8个类群,各类群材料的耐湿系数及差异如图 1所示,所有性状在各类群间的差异均具有统计学意义(p<1%,表 3). NGL中,Ⅴ类群最优;SFW中,Ⅲ类群和Ⅴ类群最优;SDW中,Ⅲ类群最优;MDA中,Ⅳ类群和Ⅴ类群最优;SPC中,Ⅱ类群最优;SSC中,Ⅶ类群最优;SOD中,Ⅷ类群最优;POD中,Ⅳ类群最优.
-
基于各性状标准化耐湿系数进行主成分分析,各主成分因子特征值、贡献率和累计贡献率如表 4所示. 当选取前6个因子时,累计贡献率为90.24%,可基本代表所有性状的全部信息,6个因子对应的特征向量值如表 5所示. 在Factor 1中,SDW,SFW和NGL所占的比例较大;在Factor 2中,SSC和POD所占的比例较大;在Factor 3中,MDA和SPC所占的比例较大;在Factor 4中,SOD所占的比例最大;在Factor 5中,POD所占的比例最大,在Factor 6中,MDA和SSC所占的比例较大.
计算各材料的主成分因子综合值,CY13PXW-17、97096、皓油11号、07191、9F087、甲972、07094、11-P74-8父本、1472和杨油6号为0.553~0.646,位于后10位;沪油15、浙双6号、01111、阳光198、CY16PXW-35、皖油15号、宁油16、史力佳、11-育7-103和CY19PXW-65为1.005~1.233,位于前10位(表 6).
-
计算各材料耐湿系数的平均隶属函数值,07191、9F087、1472、07094、97096、07022、CY13PXW-17、1321、09-P64-1和11-P74-8父本为0.199~0.246,位于后10位;阳光198、沪油14、CY20PXW-66、CY19PXW-65、SWU43、01111、CY17PXW-58、宁油1号、11-9-700和湘油15号为0.502~0.581,位于前10位(表 6).
-
计算各材料耐湿系数的综合关联度,07191、9F087、97096、1472、CY13PXW-17、07094、甲972、A148、07022和川油18为0.654~0.666,位于后10位;浙双6号、甲预25棚、阳光198、01111、宁油1号、沪油15、沪油14、CY20PXW-66、11-9-700和CY19PXW-65为0.743~0.771,位于前10位(表 6).
-
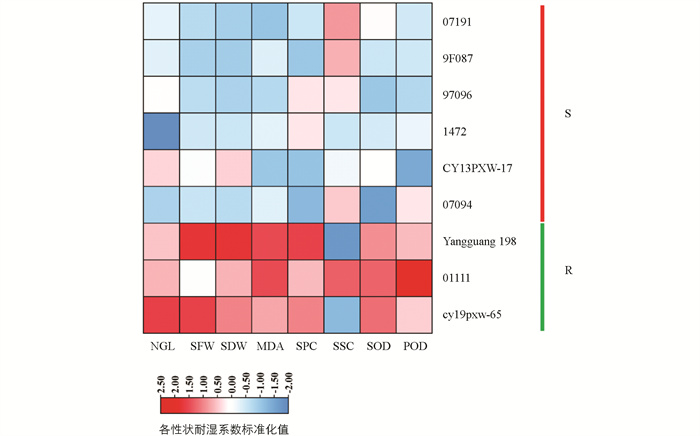
在主成分分析、隶属函数分析和灰色关联度分析中,07191、9F087、97096、1472、CY13PXW-17和07094的主成分因子综合值、平均隶属函数值和综合关联度均位于后10位,这些为湿害敏感材料;而阳光198、01111和CY19PXW-65的主成分因子综合值、平均隶属函数值和综合关联度均位于前10位,为耐湿材料. 湿害敏感材料均位于Ⅰ类群,耐湿材料位于Ⅳ类群和Ⅴ类群(表 6、图 2).
2.1. 描述统计和方差分析
2.2. 聚类分析
2.3. 主成分分析
2.4. 隶属函数分析
2.5. 灰色关联度分析
2.6. 综合评价
-
湿害胁迫下,植物会发生一系列变化,叶绿体死亡加快,单株绿叶面积减少,鲜质量和干质量降低[27-28];细胞膜发生过氧化,丙二醛等有害物质过量积累,超氧化物歧化酶和过氧化物酶等协同作用参与过氧化防御反应[29-30];大分子碳水化合物和蛋白质分解加强而合成受阻,可溶性糖和可溶性蛋白质量分数发生变化[31-32]. 本研究发现,湿害胁迫导致油菜幼苗绿叶数减少,地上部鲜质量和干质量降低,丙二醛质量分数和可溶性糖质量分数升高,过氧化物酶活性增强,可溶性蛋白质量分数和超氧化物歧化酶活性降低,这与前人研究结果基本一致[17, 19, 21, 33-35].
-
植物对环境胁迫产生的响应是多方面的,涉及生长发育、生理生化、分子机制等,单一性状难以准确地反映不同材料的胁迫耐受性. 在油菜苗期耐湿性评价性状选择方面,大多数研究者选取简单易测的生长发育形态性状,如李真等[17]和涂玉琴等[21]测定了根长、地上部干质量、根干质量和植株总干质量等;尹继春等[13]测定了绿叶数、叶面积、株干质量、根茎粗、伤流量和地上植株碳/氮等. 也有部分学者增加了部分生理生化性状评价,如陈娟妮等[19]测定了根干质量、总干质量、根系活力、叶绿素质量分数、脯氨酸质量分数和蛋白质质量分数等;薛远超[18]测定了地上部鲜质量、地上部干质量、根长、根部干质量、叶绿素质量分数、可溶性糖质量分数、可溶性蛋白质量分数、丙二醛质量分数、脯氨酸质量分数和根系活力、叶片保水力、质膜透性等,对油菜耐湿性进行了评价.
在油菜耐湿性鉴定评价中,前人多采用简易、单一的方法进行,有的直接进行性状比较,有的使用诸如活力指数[15-16]、耐湿指数[13]、综合耐湿指数[19]等,研究结果的准确性和可靠性较差;也有的使用主成分分析和二维排序分析法[20]、主成分分析结合隶属函数分析[21]进行综合评价,但这种研究目前还较少. 聚类分析、隶属函数分析、主成分分析和灰色关联度分析等方法可将多个单一性状转化为综合指标,目前已被广泛应用于环境胁迫抗性等复杂性状的综合评价中. 张春宵等[36]通过相关性分析、聚类分析和主成分分析研究了土培条件下玉米萌发期的耐旱鉴评技术体系. 霍宏亮等[37]利用主成分分析、热图和聚类分析对杜梨的耐盐碱性进行了评价,筛选出了高抗盐碱型和盐碱敏感型资源. 王薇薇等[38]利用主成分分析和隶属函数对大蒜萌发期的耐盐性进行了综合评价. 祝令晓等[39]采用主成分分析、模糊隶属函数、聚类分析、相关性分析评价了棉花各品种苗期耐低氮能力.
本研究测定了油菜幼苗绿叶数、地上部鲜质量和干质量、叶片丙二醛质量分数、超氧化物歧化酶活性、过氧化物酶活性、可溶性糖质量分数和可溶性蛋白质量分数等反映湿害胁迫对植株生长、渗透调节和过氧化防御影响的8个性状,基于各性状的耐湿系数,将系统聚类、主成分分析、隶属函数分析和灰色关联度分析4种综合评价方法相结合,对甘蓝型油菜耐湿性进行分类和综合评价,并将所有材料划分成具有显著差异的8个不同类群,进一步筛选出在各方法中均位于后10位的6份湿害敏感材料和均位于前10位的3份耐湿材料. 与已有的研究相比,本研究选取的评价性状更为合理,获得的评价结果更为可靠,且耐湿材料可在油菜耐湿性新品种培育中加以利用,亦可作为极端材料在油菜耐湿性机理研究中被采用.
-
本研究的200份甘蓝型油菜材料来源于国内外多家油菜研究单位,筛选出的湿害极端材料包含1份常规品种和8份品系. 6份湿害敏感材料来源于湖北(4份)、四川(1份)和湖南(1份),3份耐湿材料来源于湖北(2份)和四川(1份),这是否暗示着不同湿害极端材料具有不同的地域特征,还需进行深入分析. 本研究结果还表明,6份湿害敏感材料均属于Ⅰ类群,地上部鲜质量和干质量、叶片可溶性蛋白质量分数、可溶性糖质量分数、过氧化物酶活性和超氧化物歧化酶活性的耐湿性较差;耐湿材料阳光198和CY19PXW-65属于Ⅴ类群,绿叶数、地上部鲜质量和干质量、叶片丙二醛质量分数的耐湿性较好,01111属于Ⅳ类群,绿叶数、叶片丙二醛质量分数和超氧化物歧化酶活性表现较好. 由于研究材料的遗传背景和系谱来源不清楚,对湿害极端材料之间的遗传关系分析是今后相关研究应重点关注的内容之一.
3.1. 湿害对油菜苗期的影响
3.2. 耐湿性综合评价性状和方法选择
3.3. 湿害极端材料的地域特性和遗传关系
-
本研究比较了200份甘蓝型油菜种质资源苗期湿害胁迫和正常灌溉条件下生长性状和生理生化性状的差异,发现湿害胁迫使幼苗绿叶数、地上部鲜质量、地上部干质量、叶片可溶性蛋白质量分数和超氧化物歧化酶活性显著降低,叶片丙二醛质量分数、可溶性糖质量分数和过氧化物酶活性显著升高. 聚类分析将200份种质资源分为8个类群,各类群种质资源在8个性状中表现出不同的耐湿性. 基于主成分分析、隶属函数和灰色关联度方法筛选出6份湿害敏感材料和3份耐湿材料,为油菜耐湿新品种培育和耐湿机理研究提供了基础材料.
 DownLoad:
DownLoad: