





-
开放科学(资源服务)标识码(OSID):

-
风味被定义为味道和气味的组合[1]. 植物能够合成成千上万种具有不同味道和气味的初级和次级代谢产物,蔬菜的主要代谢产物包括糖、酸、盐化合物和挥发性化合物. 蔬菜风味品质的评价指标主要包括糖、酸和挥发性化合物(醇类、醛类、酮类、萜类、脂类和含硫化合物等)的含量及组合[2-4]. 挥发性化合物主要以脂肪酸、氨基酸、色素、羟基酸等为合成前体[5],不同挥发性化合物的含量及组合决定了各种蔬菜所具有的独特风味和风味品质[5]. 蔬菜的营养和风味品质主要受作物的遗传因素制约,同时也受栽培环境、种植技术和采后贮藏处理等因素影响[6]. 近年来,随着生物化学技术、分子生物学技术突飞猛进地发展,全基因组、转录组、蛋白质组和代谢组的数据组成了生物学大数据,挥发性化合物的合成途径及其过程中相关关键酶不断被发现[7]. 确定关键酶及其控制基因,通过关键酶及其基因的过量表达或下调,利用基因DNA重组、转化等技术改善蔬菜风味品质已经成为可能[8].
我国土壤全硼质量分数在0~500 mg/kg之间,平均质量分数为64 mg/kg,大致呈由北向南、自西向东逐渐降低的趋势[9]. 当土壤有效硼质量分数低于0.5 mg/kg时,作物就会出现缺硼症状,低于0.25 mg/kg时则会出现严重缺硼症状. 全国第二次土壤普查数据表明,我国耕地土壤缺硼的面积多达3 300万hm2,西南、华南地区各省份,华北、东北地区部分省份以及长江中下游地区的耕地缺硼比例均大于60%[10]. 硼供应量不足使植物生长缓慢,植株呈丛生状,节间缩短,生长停止;花粉数量减少,受精不良;果实生长发育受阻,结实少甚至不结实. 此外,硼缺乏还会抑制植物的光合作用能力,改变植物酚类代谢,并影响氮的吸收和同化进而影响作物的风味品质[11].
大白菜(Brassica pekinensis (Lour.) Rupr.)是原产于我国的一种芸薹属叶用蔬菜,在世界各地均有种植,因其适口性好,口味佳,深受人们喜爱. 十字花科蔬菜(包括花椰菜、甘蓝和白菜)对硼的需求量很高,并且对低硼胁迫很敏感,常出现硼缺乏症[12]. 目前,国内外已对增施硼肥改善作物营养品质做了大量研究,但对硼影响蔬菜风味品质的研究依旧较为缺乏,其机制和原理也需要得到更多探索和论证. 为此本研究采用盆栽试验,通过研究不同硼肥用量对大白菜生长发育、风味品质以及谷氨酰胺合成酶(GLN)家族基因表达的影响,初步探讨了硼对大白菜风味品质的调控机理.
HTML
-
栽培基质以草炭、珍珠岩(粒径3~6mm,购自北碚花市)按体积比例3∶1混合,混合基质的基本理化性质为pH值6.30,容质量0.161 g/cm3,总孔隙度94.41%,持水孔隙75.59%,有机质213.2 g/kg,全氮3.24 g/kg,碱解氮0.62 g/kg,有效磷16.18 mg/kg,速效钾188.46 mg/kg,有效硼0.394 mg/kg. 试验蔬菜为大白菜,品种为‘华良早5号’和‘脆甜白2号’(北碚区农户大量种植品种),于重庆市北碚区三圣镇德圣村(106°38′25.77″N,29°54′53.84″E)育苗35 d.
-
试验于2018年3月20日—2018年5月12日在西南大学1号温室大棚内进行,采用基质盆栽试验. 试验采用顶部直径27 cm、底径19 cm、高21 cm的无孔灰色不透光塑料桶作为盆栽桶,每桶装填混合基质4.4 L. 试验设置5个处理: 不含硼营养液(CK)、含0.5 mg/L H3BO3营养液(B1)、含1 mg/L H3BO3营养液(B2)、含2 mg/L H3BO3营养液(B3)、含4 mg/L H3BO3营养液(B4),每个处理设置4次重复,随机排列. 3月20日进行大白菜定植工作,选取长势一致、健壮的幼苗(四叶一心)移栽,每盆3株,移栽时浇500 mL的1/2无硼日本园试营养液(表 1),之后每3 d浇300 mL营养液以补充养分. 4月4日进行不同硼梯度处理,每3 d浇300 mL的不同硼质量浓度的营养液,观察并记录其生长状况,5月12日收获. 整个生长期间浇灌营养液后使用去离子水补充水分,保持基质含水量为田间持水量的70%~80%. 培养50 d收获.
-
基质pH值采用土水比1∶2.5混合,雷磁pH计(PHSJ-5,China)测定;栽培基质容质量、总孔隙度、孔隙度采用环刀法测定[13];将环刀浸入水中至栽培基质饱和,然后用烘干法测定饱和含水率[14];采用常规方法测定栽培基质全氮、碱解氮、有效磷、速效钾、有效硼、有机质等理化性质[15].
-
氨基酸组分参照GB 5009.124-2016《食品安全国家标准食品中氨基酸的测定》,使用日立L-8800型氨基酸自动分析仪测定.
-
RNA提取采用生工生物工程(上海)股份有限公司EZ-10 DNAaway RNA Mini-Preps Kit试剂盒. RNA提取后使用Nano Drop 2000C分光光度计检测RNA浓度,通过OD260/280和OD260/OD230的比值结合1%琼脂糖凝胶电泳检测RNA的降解程度和质量,提取合格的RNA样品置于-80 ℃超低温冰箱保存.
-
使用TaKaRaPrimeScriptTM Reagent Kit with Gdna Eraser(Perfet Real Time)试剂盒对提取合格的RNA进行残存基因组DNA去除和逆转录处理,cDNA置于-20 ℃保存备用.
-
本试验研究了与氮代谢相关的谷氨酰胺合成酶(Glutamine Synthetase,GS)同工型GLN家族基因. 在拟南芥(Arabidopsis thaliana L.)数据库中获取拟南芥GLN家族基因的基因座编号,在NCBI网站(
https://www.ncbi.nlm.nih.gov/ )进行BLAST处理,在Vector NTI Advance 11.5上进行拟南芥与大白菜GLN家族基因序列对比,在Primer 5.0/OligoArchitecxtTMOnline(http://www.oligoarchitect.com )上进行设计工作,共设计4对qRT-PCR引物(表 2). 设计引物由重庆擎科兴业生物技术有限公司合成. -
使用PCR仪(ABI-9700,America)对cDNA进行特异扩增. PCR反应体系为: 2.5 μL 10×Easy Taq Buffer (Mg2+),0.35 μL EasyTaq DNA Polymerase,0.5 μL dNTPs,0.5 μL正向引物F (5'-GTTACCACAGGGATAACTGGCTTG-3'),0.5 μL反向引物R(5'-CTAACCTGTCTCACGACGGTCTAA-3'),20.15 μL dd H2O,0.5 μL cDNA. PCR反应程序为: 94 ℃预变性2 min,98 ℃变性30 s,退火(退火温度为60.0 ℃) 30 s,72 ℃延伸30 s,35个循环,72 ℃最后延伸10 min,16 ℃ 3 min.
-
分别以大白菜各处理混合cDNA为模板,以60 ℃为基准,设置58,60,62,64,65,66 ℃ 6个温度梯度,采用25 μL标准半定量RT-PCR体系,在不同温度下扩增各样品中GLN家族基因,PCR反应体系为: 0.5 μL正向引物F,0.5 μL反向引物R,12.5 μL FastStart Essential DNA Green Master,11 μL dd H2O,0.5 μL cDNA. PCR反应程序为: 95 ℃预变性10 min,95 ℃变性10s,退火(退火温度为设置的6个温度梯度) 30 s,72 ℃延伸20 s,45个循环,添加熔解曲线65 ℃到95 ℃. 扩增产物进行1%的琼脂糖凝胶电泳,以检测各引物的最适退火温度.
-
以2个品种大白菜5个不同处理的反转录产物cDNA稀释30倍后作为模板,26SrRNA为内参基因,使用荧光定量PCR仪(CFX96TM Real-Time System)进行qRT-PCR检测,反应体系为: 5 μL Fast Start Essential DNA Green Master,0.5 μL正向引物F,0.5 μL反向引物R,1.5 μL dd H2O,2.5 μL cDNA. qRT-PCR反应程序为: 95 ℃预变性10 min,95 ℃变性10 s,T1温度(T1为不同引物的最适退火温度)退火10 s,72 ℃延伸20 s,45个循环,添加熔解曲线65 ℃到95 ℃.
-
利用Microsoft Excel 2016进行数据处理和图表制作,IBM SPSS Statistics 21.0进行主成分分析、单因素方差分析(One-way ANOVA)、相关性分析(Pearson相关系数法)、最小显著差异法(Fisher Least Significant Difference,LSD)检测法进行不同处理均值的差异显著性比较,当p<0.05时,差异有统计学意义.
1.1. 试验材料
1.2. 试验设计
1.3. 测定指标及方法
1.3.1. 基质理化性质
1.3.2. 氨基酸组分
1.3.3. RNA提取与质量检测
1.3.4. RNA纯化和逆转录
1.3.5. GLN家族基因引物设计
1.3.6. 逆转录产物cDNA扩增
1.3.7. 引物退火温度梯度试验
1.3.8. GLN家族基因qRT-PCR检测
1.4. 数据处理与统计分析
-
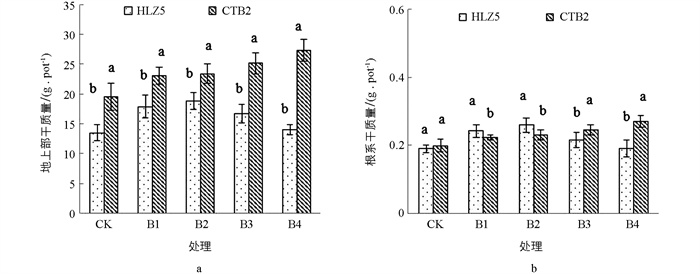
不同硼质量浓度处理(0,0.5,1,2,4 mg/L H3BO3)下不同品种大白菜的地上部、根系生物量如图 1所示. 施用硼肥后,‘华良早5号’的地上部、根系随着硼质量浓度的提高呈先增加后减少的趋势(在最高硼质量浓度处理下仍高于对照),与对照组相比,各处理的地上部和根系干质量分别增加3.32%~39.66%和0.48%~36.93%,在1 mg/L H3BO3处理下各生物量最大;‘脆甜白2号’的地上部和根系干质量随着硼质量浓度的提高而增加,各处理的地上部、根系较对照组分别增加18.16%~40.08%和12.12%~36.51%,在4 mg/L H3BO3处理时达到最大值. 此外,不同大白菜品种间对高硼的耐受能力存在差异,本试验中‘脆甜白2号’对高硼的耐受能力强于‘华良早5号’.
-
如图 2所示,大白菜根部GLN家族基因的表达量随硼质量浓度的提高表现出不同的变化趋势: 外源施用硼肥降低了大白菜根部GLN1.1的表达量;‘华良早5号’根部GLN1.2的表达量表现为先上调后下调最后再上调,‘脆甜白2号’则表现为先下调再上调,2个品种大白菜GLN1.2的表达量均在B4处理下最大,分别较对照上调54.69倍和181.18倍;‘华良早5号’根部GLN1.4的表达量较对照表现为下调,‘脆甜白2号’表现为先上调后下调,B1,B2和B4处理分别较对照上调1.04,1.14和1.1倍;GLN2的表达量在‘华良早5号’中仅在B3处理上调1.65倍,在‘脆甜白2号’表现为先上调后下调,B1,B2和B3处理分别较对照上调20.31,1.51,38.61倍.

如图 3所示,大白菜叶柄GLN家族基因的表达量随硼质量浓度的提高表现为: ‘华良早5号’叶柄GLN1.2和GLN2的表达量随硼质量浓度的提高表现为先下调后上调,B3处理较对照分别上调7.06倍和3.09倍;GLN1.1和GLN1.4的表达量则不断上调,分别在B4和B3处理下达到最大值,较对照分别上调7.66倍和131.97倍;‘脆甜白2号’叶柄GLN1.1的表达量除在B3处理中下调外,B1,B2和B4处理分别较对照上调1.43,2.13,3.14倍;GLN1.4和GLN2的表达量表现为先上调后下调,均在B2处理下达到最大值,分别较对照上调1.02倍和322.85倍;GLN1.2的表达量则在外源施用硼肥后表现为不断下调.
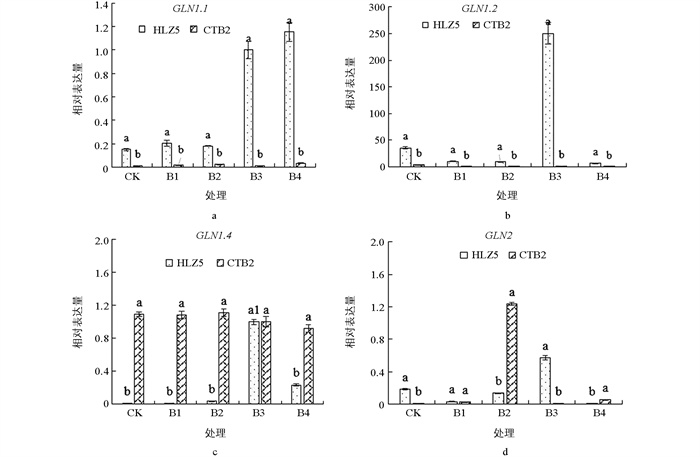
如图 4所示,大白菜叶片中GLN家族基因的表达量在不同质量浓度硼酸处理下表现趋势为: GLN1.1表现为先下调后上调,‘华良早5号’和‘脆甜白2号’B1,B2,B3,B4处理下的表达量分别为对照的0.46,92.37,23.32,24.14倍和0.6,1.74,4.01,6.96倍;‘华良早5号’ GLN1.2的表达量表现为先下调后上调,各处理的表达量分别为对照的0.86,59.78,61.17,94.86倍;‘脆甜白2号’ GLN1.1的表达量在B1和B3处理下上调,各处理分别是对照的1.60,0.30,2.14,0.55倍;‘华良早5号’ GLN1.4的表达量表现为先上调后下调,‘脆甜白2号’则为先下调后上调,2个品种大白菜GLN1.4的表达量分别在B2和B3处理下达到最大值,较对照分别上调7.29倍和21.23倍;‘华良早5号’和‘脆甜白2号’各处理下GLN2的表达量均较对照上调,分别较对照上调1.36,278.62,398.44,93.77倍和1.07,1.73,1.77,1.52倍.

-
大白菜氨基酸组成及其质量分数随着硼质量浓度和品种的变化如表 3所示. ‘华良早5号’的必需氨基酸、半必需氨基酸和氨基酸总量随着硼质量浓度的提高呈先上升再降低的趋势,均在B2处理达到最大值,分别较对照增加23.74%,27.38%和20.21%;均在B4处理达到最小值,分别是对照的85.25%,68.72%和83.84%;外源补充硼元素提高了EAA/TAA,从而提高了大白菜的营养价值. ‘脆甜白2号’的必需氨基酸、半必需氨基酸和氨基酸总量随着硼质量浓度的提高表现出和‘华良早5号’相反的趋势,即先降低再上升;半必需氨基酸质量分数分别在B4,B3处理下达到最大、最小值,必需氨基酸和氨基酸总量均在对照、B2处理取得最大、最小值;EAA/TAA在低硼质量浓度处理下(B1,B2)稍低于对照组,在较高硼质量浓度处理下(B3,B4)高于对照组.
-
蔬菜氨基酸的组成及质量分数不尽相同,也有众多评价蔬菜氨基酸营养价值的方法,但蔬菜所含必需氨基酸的种类、数量和组成比例等始终是决定蔬菜营养价值高低的重要指标. 本试验采用了主成分分析法(PCA)分析不同硼酸质量浓度处理对大白菜氨基酸营养价值的影响. 如表 4所示,利用主成分分析法(提取条件为: 特征值>1,累计方差贡献率≥85%)在大白菜17种氨基酸中提取出3个主成分. 主成分1,2,3的特征值和方差贡献率分别为11.387,2.964,1.162和56.59%,25.74%,8.93%,累计方差贡献率为91.25%,可以代表大白菜氨基酸营养价值的大部分信息.
表 5是大白菜17种氨基酸的荷载量,各个变量在主成分中的重要程度通过数值的高低表现出来. PC1包括了大白菜氨基酸营养价值的绝大部分品质信息,其中天门冬氨酸(Asp)、谷氨酸(Glu)、甘氨酸(Gly)、异亮氨酸(Ile)和赖氨酸(Lys)系数较大,因此将以上5种氨基酸作为第1主成分的代表指标. PC2则是苏氨酸(Thr)和脯氨酸(Pro)系数较大,所以把苏氨酸和脯氨酸作为第2主成分的代表指标. PC3主要反映了大白菜甲硫氨酸(Met)贡献最大,故第3主成分的代表指标为甲硫氨酸.
为了构建大白菜氨基酸3个主成分模型,把17种氨基酸的主成分因子载荷除以其对应特征值的算术平方根,即得到每种氨基酸变量Xkn的系数Ykn,以此为权重得到主成分的表达式如下:
式中: Fk为主成分k的得分;Ykn为变量Xkn的系数;Xkn为主成分k的第n个因子载荷.
以3个主成分各自对应特征值的权重建立综合得分评价模型,表达式为F=56.59/91.251×F1+25.735/91.251×F2+8.925/91.251×F3. 由此计算2个品种大白菜在不同处理下氨基酸营养价值的得分及排名,如表 6所示. 根据综合得分值,‘华良早5号’各处理的氨基酸营养价值由高到低为: B2,B3,CK,B1,B4,‘脆甜白2号’各处理由高到低为: CK,B4,B1,B2,B3,‘华良早5号’的氨基酸营养价值总体上高于‘脆甜白2号’.
-
蔬菜中氨基酸可分为游离氨基酸和非游离氨基酸,其中游离氨基酸对食物风味的影响较大. 按照氨基酸味觉的差别,一般把氨基酸分为甜味氨基酸(Pro,His,Ala,Gly,Ser,Thr)、苦味氨基酸(Val,Met,Ile,Leu,Arg)、鲜味氨基酸(Lys,Asp,Glu)和芳香族氨基酸(Cys,Tyr,Phe),各风味氨基酸的相对质量分数动态变化并相互影响,因此形成了各类食物多种多样的风味. 由表 7可知,‘华良早5号’地上部味觉氨基酸质量分数从高到依次为: 甜味氨基酸、苦味氨基酸、鲜味氨基酸、芳香族氨基酸,‘脆甜白2号’则为: 甜味氨基酸、鲜味氨基酸、苦味氨基酸、芳香族氨基酸;2个品种大白菜甜味氨基酸、苦味氨基酸、鲜味氨基酸和芳香族氨基酸分别占味觉氨基酸总量的31.78%~34.03%,28.99%~30.71%,26.49%~28.07%,9.14%~9.96%;30.61%~33.83%,28.21%~29.04%,28.62%~30.26%,9.34%~10.47%. 外源添加硼增加了苦味氨基酸和芳香族氨基酸的相对含量,减少了甜味氨基酸的相对含量.
由于不同氨基酸的味觉阈值[16]存在较大差异,有的甚至相差了几个数量级,通过味觉氨基酸的含量来评价各种氨基酸对食物风味品质贡献的方法是不严谨的,因此本试验采用大白菜中各种味觉氨基酸含量与其味觉阈值之比(Ratio of Content and Taste Threshold,RCT)实现这一目标. 一般认为,RCT值≥1的氨基酸对食物的整体风味有较大贡献,且比值和影响成正比;RCT值<1的氨基酸则被认为对食物的整体风味无影响. 如表 8所示,外源硼元素的加入对大白菜各氨基酸RCT产生了不同的影响. 对大白菜风味贡献最大的天门冬氨酸和谷氨酸在‘华良早5号’中先增后减,B2处理达到最大值;‘脆甜白2号’则是先减后增,对照处理达到最大值. 总之,‘华良早5号’不同硼质量浓度处理下味觉氨基酸RCT值由大到小依次为: B2,B3,B1,CK,B4,‘脆甜白2号’则为: CK,B4,B1,B3,B2. ‘华良早5号’中对风味贡献较大的氨基酸的含量阈值比总体大于‘脆甜白2号’,‘华良早5号’较‘脆甜白2号’可能具有更好的风味品质.
-
大白菜地上部17种氨基酸含量与GLN家族基因表达量的相关性分析结果如表 9所示. 大白菜地上部苏氨酸(Thr)含量与根部GLN1.1和GLN1.4显著正相关(r=0.658*,r=0.694*),半胱氨酸(Cys)含量与叶柄GLN1.4极显著正相关(r=0.817*),酪氨酸(Tyr)含量与叶片GLN1.4显著负相关(r=-0.703*),组氨酸(His)含量与叶柄GLN1.4显著正相关(r=0.676*),脯氨酸(Pro)含量与根部GLN1.1,GLN1.4的相关性分别达到极显著和显著水平(r=0.772**,r=0.651*),其他氨基酸和GLN家族基因之间的相关性均未达到显著或极显著水平.
2.1. 不同硼肥用量对大白菜生物量的影响
2.2. GLN家族表达量分析
2.3. 不同硼肥用量对大白菜氨基酸组成及质量分数的影响
2.3.1. 氨基酸组成和质量分数
2.3.2. 氨基酸营养价值分析
2.3.3. 氨基酸风味品质分析
2.4. GLN家族基因表达量与氨基酸的相关性分析
-
硼与植物生长的变化、固氮和硝化作用、次级代谢和氧化应激等密切相关[17]. 本试验中,硼缺乏(CK)显著抑制了大白菜植株的生长,通过外源补充硼肥后,大白菜根部、地上部生物量随着硼质量浓度的提高而显著增大,‘华良早5号’和‘脆甜白2号’的根部和地上部生物量分别在B2和B4处理达到最大值,株高分别在B3和B2处理达到最大值,但随后生物量开始减少. 原因可能是硼缺乏和产生毒害之间的范围很小,当外源添加硼元素使基质中硼的质量浓度超过这一范围后,就对植株的生长发育产生不利影响甚至产生毒害作用. 同时,2个品种大白菜的生物量对硼的响应不完全一致,可能是2个品种对硼需求量的差异导致的,‘脆甜白2号’对硼的需求量大于‘华良早5号’.
氨基酸既是蔬菜营养品质的重要组成部分,又是影响蔬菜风味的重要因素之一[18]. 本试验中,施用硼肥提高了‘华良早5号’EAA/TAA,必需氨基酸和非必需氨基酸达均在B2处理下达到最大值,‘脆甜白2号’在B3,B4处理下的必需氨基酸、非必需氨基酸和EAA/TAA均高于对照,说明适当质量浓度的硼可以提高大白菜氨基酸质量分数,提升大白菜的营养价值. 本试验对不同硼酸处理下大白菜地上部氨基酸的营养价值进行主成分分析法时,综合考虑了氨基酸的总量和组成,‘华良早5号’和‘脆甜白2号’的氨基酸营养价值分别在B2和B4处理最大,‘华良早5号’较‘脆甜白2号’有更高的氨基酸营养价值. 在对大白菜地上部的氨基酸风味品质分析中发现,甜味氨基酸质量分数最高,是大白菜的主要呈味氨基酸. 此外,我们还引用了味觉阈值来分析不同处理下大白菜地上部氨基酸的风味品质,发现鲜味氨基酸中的天门冬氨酸和谷氨酸对大白菜风味贡献最大,‘华良早5号’和‘脆甜白2号’不同硼质量浓度下味觉氨基酸RCT值由大到小依次为: B2,B3,B1,CK,B4;CK,B4,B1,B3,B2,‘华良早5号’较‘脆甜白2号’有更高的氨基酸风味品质. 适当施用硼肥可以提高大白菜地上部氨基酸营养价值和风味品质,可能是硼对氮代谢产生促进作用,使植物体内与氮代谢的相关酶(如硝酸还原酶、谷氨酰胺合成酶和天冬酰胺合成酶)的活性增加[19],促进了植物体内氨基酸的合成.
目前已有大量研究表明植物体内的硼氮具有相互促进的作用[20]. 根部吸收硝酸盐被认为是氮代谢的关键过程,虽然目前尚无令人信服的证据表明硼对硝酸盐的同化有直接作用[21],但Camacho-Cristóbal等[22]的研究显示,缺硼条件下烟草叶片和根部的硝酸盐质量浓度均下降,表明维管植物中硼与硝酸盐同化之间可能存在联系. 硝酸盐(NO3-)和铵盐(NH4+)是植物根部吸收无机氮的主要形式[23],植物吸收的无机氮转化为NH4+后被同化到有机氮化合物中,随后掺入蛋白质、核酸和一系列次级代谢产物中[24]. “GS-GOGAT循环”是植物氨同化最基本的途径,GS在“GS-GOGAT循环”中作为催化氨初始同化的关键酶,起到氨库的作用[25]. 目前在高等植物的基因组中编码了两种GS同工型: 胞质Gln合成酶同工型(GS1/GLN1)和叶绿素Gln合成酶同工型(GS2/GLN2). 由于GS1和GS2的编码差异,以及它们分布在植物不同器官的特异性,决定了它们具有不同的生理作用[26]. 在大多数植物物种中,GS在叶绿体(GS2/GLN2)中以单一同工型的形式存在,在细胞质中以5种同工型的形式存在[27]. GS2的主要作用是同化NO3-还原和光呼吸释放的NH4+;GS1的主要作用是产生谷氨酰胺以完成细胞之间氮的运输,但单个GS1亚型可能发挥不同的功能. 大麦(Hordeum vulgare L.)基因组编码3种GS1同工酶,其中HvGS1-1转录本主要存在于维管组织中,HvGS1-2主要存在于根部,而HvGS1-3几乎仅存在于籽粒中[28]. 数量性状基因座的研究表明,大麦HvGS1-1基因与谷物蛋白质含量有关[29],而玉米直向同源物ZmGln1-3和ZmGln1-4分别与谷粒质量和籽粒产量相关[30]. 硬粒小麦(Triticum durum L.)中籽粒蛋白质含量、GS(GS1和GS2)基因表达与GS活性之间呈正相关[31-32].
相对于谷氨酸和铵,不同的GS1亚型表现出不同的动力学性质: GLN1.1和GLN1.4对铵具有高亲和力,而GLN1.2和GLN1.3具有低亲和力,GLN1基因在根、茎和叶片衰老过程中差异表达[33]. 在拟南芥的6个GLN基因中,GLN1.1,GLN1.2,GLN1.3,GLN1.4在根中高度表达[34],氮缺乏导致根中GLN1.1,GLN1.3和GLN1.4上调[24]. 本试验中,除‘脆甜白2号’叶柄的GLN1.4外,施用硼肥提高了大白菜叶柄和叶片GLN家族基因的表达量,可能是因为施用硼肥后增强了大白菜根系对无机氮(以NO3-为主)的吸收能力[22],NO3-通过维管束运输到叶片后被转化为NH4+,从而提高了叶片NH4+的水平,提高了叶柄和叶片GLN家族基因的表达量[35],但是大白菜根GLN1.1和GLN1.4的表达量随着硼质量浓度的提高而下调. 大白菜不同器官的GLN家族基因表达量对不同质量浓度硼的响应存在差异可能是由于GLN表达过程的复杂性、作物品种的差异性,以及氮素对GLN的影响具有细胞特异性导致的[36],其具体的作用机制还需要进一步研究. 此外,本试验使用Pearson相关性分析方法,对不同硼质量浓度处理下大白菜GLN基因家族表达量分别与氨基酸和挥发性化合物进行相关性分析,发现前述部分物质与GLN基因家族的相关性达到显著或极显著水平,其机理还需进一步深入探讨.
-
在缺硼的条件下适当补充硼肥可以提高大白菜的生物量. 整体上看,B2 (1 mg/L H3BO3)处理下大白菜生长情况最好. 不同大白菜品种间对高硼的耐受能力存在差异,本试验中,‘脆甜白2号’对高硼的耐受能力强于‘华良早5号’. 施用硼肥提高了大白菜必需氨基酸占氨基酸总量的比例. 蛋氨酸和半胱氨酸是大白菜的第一限制氨基酸,鲜味氨基酸中的天门冬氨酸和谷氨酸对大白菜风味贡献最大. 施用硼肥对大白菜根、叶柄和叶片GLN家族基因表达量产生不同的影响,叶片GLN家族基因表达量总体上调. 大白菜地上部半胱氨酸质量分数与叶柄GLN1.4极显著正相关,酪氨酸质量分数与叶片GLN1.4显著负相关,组氨酸质量分数与叶柄GLN1.4显著正相关.
 DownLoad:
DownLoad: