





-
开放科学(资源服务)标识码(OSID):

-
湿地是地球上最丰富的生态系统之一,提供了重要的生态服务,如水质净化、洪水调节和栖息地供给等[1]. 然而,由于人类活动的干扰和外来物种的引入,湿地植物群落面临着许多威胁[2-3]. 外来植物物种的引入可能是有意的,例如用于园艺等农业目的;也可能是无意的,例如通过船只或风传播的种子. 无论是有意还是无意引入,外来植物物种都有可能对湿地植物群落造成不利影响[4]. 既可能对湿地植物群落的物种组成和多样性产生影响,如竞争湿地乡土植物的生存空间、养分和光线资源,导致乡土植物数量减少或消失[5];也可能导致湿地植物群落的物种多样性降低,从而降低湿地生态系统的稳定性,还可能破坏湿地的水质净化和洪水调节功能,导致水质恶化和洪水风险增加[6]. 因此,研究人类引进外来植物物种对湿地植物群落的影响对湿地生态系统的保护和管理至关重要.
人类引进外来植物物种使植物群落产生了广泛而复杂的变化,这些外来物种可以改变当地物种的组成、结构和功能,对生态系统产生重要的影响,且外来植物在河岸带的引入、传播和影响被广泛认为对生态系统起到了重要的推动作用[7]. 外来植物的引入主要是由人类活动所致,例如引进外来植物作为观赏植物、农作物或园艺植物,或者是通过人类活动扩张和改变自然生境,使外来植物有机会进入并传播到河岸带. 这些外来植物的传播能力和适应力强,往往能够迅速占领河岸带的生境,对原有植被群落产生重要的影响. 人类活动对自然生境的干扰导致河岸带植被的物种组成发生明显的变化[8]. 由于人类活动的干扰,河岸带原始植被群落逐渐被破坏,乡土植物种类减少,而外来植物种类增多. 这种物种组成的变化降低了河岸带植被群落的多样性和功能,影响了水陆生态系统的物质能量交换等生态系统功能. 因此,研究在不同人为干扰下河岸带外来植物群落的特征,对揭示快速城市化背景下河岸带植被结构的变化具有重要意义.
虽然有学者已经对河岸带植被的分布格局、物种多样性、功能群划分和生态服务功能等方面进行了研究[9],但对河道附近湿地外来植物的研究却较少. 特别是在不同人类活动干扰类型下,对外来植物群落特征和影响程度的相关研究还未见报道. 本研究选择汉江流域的一个主要支流,即河南省南阳市唐河县段的唐河湿地外来植物群落作为研究对象,通过对湿地植被进行系统调查,分析不同人类活动干扰类型下的湿地外来草本植物群落特征,以期反映社会经济不断发展背景下河道湿地外来植物群落的变化规律,为湿地生态恢复提供科学依据.
HTML
-
唐河为长江流域汉水水系的支流. 唐河经由河南的社旗、唐河、新野以及湖北襄阳市,在两河口与白河汇合后更名为唐白河,终南至张家湾,为汉水提供补给. 唐河全程230 km,流域总面积为8 685 km2,拥有众多支流. 左岸分布有毗河、泌阳河、三家河、清水河、蓼阳河、礓石河等,右岸则有桐河、绵延河、涧河等,其中流域面积超过100 km2的支流有26条.
本研究区域在唐河流域的唐河县段. 唐河县位于豫西南南阳盆地东部,与桐柏、泌阳相连,西通新野、南阳,北侧与社旗接壤,南部与湖北的枣阳相邻. 地理位置处于北亚热带与暖温带的过渡区,具有北亚热带季风型大陆气候特征. 年平均日照总时数约为2 187.8 h,年平均太阳总辐射量为116.56 kcal/cm2. 年平均气温为15.2 ℃,月平均气温在1.4~28.0 ℃范围内变化. 该地无霜期长达233 d,年平均降雨量为910.11 mm,主要降雨期集中在4-9月,降水量达689.2 mm,约占全年降水量的75.7%.
-
本研究以河南省唐河县的唐河河段为研究区域,采用完全随机试验设计开展不同区域与植物物种来源2个因素试验.
其中不同区域(C)设置2个水平,分别为乡村段唐河湿地(C1)、城市段唐河湿地(C2);植物物种来源(P)设置3个水平,分别为乡土植物(P1)、外来植物(P2)、入侵植物(P3). 试验设计见表 1.
-
2023年1-6月,对河南唐河县唐河河段进行草本植物群落土壤生态考察. 首先,河岸全部区域进行物种调研,标定出每块地带的特殊植被. 随后,根据每种植物群落的特殊类型,在每个样地设定一条样线. 这条样线遵循垂直于河流方向的设计原则,起始于河滩,每隔20 m划定一个样地,一直延伸到河岸的边界. 通过众多样品的获取,本研究得以揭示植物群落的基本属性. 在唐河县段的河岸区域,我们总共设立了8个考察点,获取了80个样本,包括4个农村段的样地(共40个样地)和4个城区段的样地(共40个样地).
该地区主要植被为针叶、阔叶混交林,典型的种类包括常绿阔叶林、落叶阔叶林、针阔混交林、常绿落叶阔叶林及常绿针叶林. 本次考察积累了每个样地草本植物群落的重要数据. 这些数据包括样地中草本植物的名称、生活习性、水源类型、个体数量、平均高度和地表生物量. 在测算个体数量时,如果存在簇生或簇状植物,每种都独立计算. 若样地只包含一类植物且分布均匀,那么将在50×50 cm2的试验区域考察样地的总体数量. 若植物种群分布不均,则会取150×150 cm2样本的地表生物量进行收集并烘干测量.
-
1) 乡土植物:本研究中乡土植物不包括本地与外来植物杂交形成的种类,而是根据植物种类在流域内自然分布及人为引入和种植的历史来确定.
2) 外来植物:进一步将外来植物分为国内外来物种和国外外来物种,通过考察其本地寄生地、人为引入的记录以及在全国范围内的主要分布地来进行判断.
3) 入侵植物:通常在其天然生长环境以外产生大量个体,并具备广泛扩散的可能性.
-
为了评估河滩带草本植物群落中各物种的优势程度,本研究使用了合成优势度指标进行计算,并使用了Sorensen相似性指数表征植物群落的相似性. 优势度和相似性分析具体计算见文献[10].
-
采用Excel 2010和SPSS 22.0软件对本文中所用到的数据进行统计分析.
2.1. 试验设计
2.2. 植被调查
2.3. 数据分析
2.3.1. 判断植物来源的方式
2.3.2. 确定优势度和相似性分析
2.4. 数据处理
-
本研究选择唐河县的唐河河段作为调查对象,发现该地区植物群落非常丰富,共包括93科维管植物,意味着在该地区存在多样性的植物群落,涵盖了不同的科属植物. 在这93科维管植物中,共有294个属和386个种类. 这种多样性在生态系统中起着重要作用,维持着植物间的相互作用和生态平衡. 在唐河河段的植物群落中,有一部分是外来植物,占所有植物的31.28%. 在外来植物中,来自国内的有18科、22属和14种,这些植物可能是由于人类活动引入或者自然扩散到了该地区. 来自国外的有27种,分布在3个科、13个属中. 这些外来物种主要来自美洲、欧洲和亚洲. 进一步分析外来物种的比例,发现这些外来植物中美洲物种占24.54%,欧洲物种占23.02%,亚洲物种占52.44%,说明唐河河段外来植物主要来自这3个地理区域,其中亚洲物种的占比最高. 在所有的外来植物中,有一些科的外来物种比例较高,例如菊科、禾本科、豆科和苋科是其中比较典型的科,特别是菊科占外来物种的比例最大.
-
由图 1可知,各区域河岸植物群体的种类构成存在显著变化(p<0.05). 乡村段乡土植物的比例最大,占总种类构成的65.38%,意味着乡村地区的河岸植物群落主要由本地的乡土植物组成,这些植物可能具有在该地区长期生存和繁衍的优势. 其次是入侵植物,占总种类构成的比例较大,表明乡村地区的河岸植物群落中也存在一定数量的入侵植物物种,这些植物可能是由于人类活动引入或者随着环境变化进入了该地区. 入侵植物的存在可能对乡土植物群落造成竞争和生态影响. 相比之下,城市河段的入侵植物比例明显少于外来植物的比例,表明城市地区的河岸植物群落中外来植物的种类更多,而入侵植物相对较少,这可能是由于城市地区人为干扰和管理措施对入侵植物的管控和抑制起到了一定的作用. 在乡土植物方面,乡村地区的数量优于城市地区,说明乡村地区相对保留了更多乡土植物的物种多样性,也保护了本地生态系统的特色和稳定性. 对于入侵植物而言,乡村地区的数量明显低于城市地区,表明随着人为干扰增加,外来植物进入乡村地区的数量明显增加,在城市地区却更容易受到管理和控制. 乡村地区和城市地区各自独有的乡土植物和外来植物种类存在差异. 乡村地区的乡土植物种类较多,有15种,而城市地区只有3种. 在外来植物方面,乡村地区有7种独有外来植物,城市地区有13种独有外来植物,说明乡村地区和城市地区在物种构成和物种多样性方面具有差异. 这些差异可能与人为干扰、管理措施、环境变化等因素密切相关,因此对植物群落的结构和生态影响值得进一步研究.

-
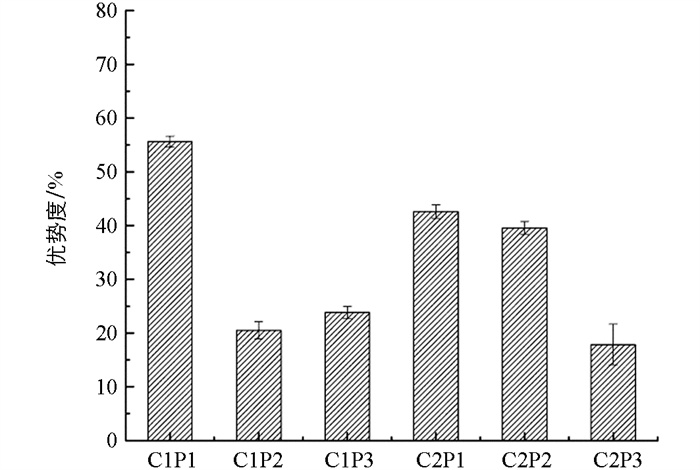
由图 2可知,随着人为干扰强度逐步提升,不同河段带植物种类的主导地位也呈现出明显的差异(p<0.05). 在本研究中农村段入侵植物形成主导地位,城市段次之. 无论农村段还是城市段,乡土植物的主导地位始终最高,其中农村段乡土植物的主导地位超过了城市段. 农村段外来植物的主导地位明显低于城市段. 同时,不同河段外来植物中的优势种出现了变化. 例如农村段的月季和城市段的白玉兰、紫荆等,都是引进的外来种.
-
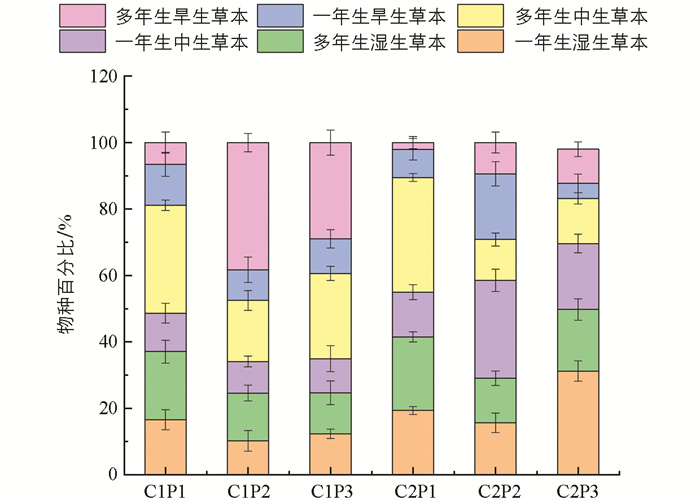
通过图 3分析可知,不同河段植物群落生活型构造存在显著差异性. 乡村地区的乡土植物群落主要由多年生中生草本植物组成,占总生活型的20.52%,意味着乡村地区的乡土植物倾向于较长生命周期的中生型草本植物,可能具有适应当地环境长期生存的特点. 在外来植物中,多年生的旱生草本植物是最常见的生活型(占38.29%),表明在乡村地区外来植物主要是适应干旱环境的多年生草本植物,可能是由于乡村地区的气候和土壤条件更适合这类植物的生存与繁殖. 入侵植物种类最丰富的生活型仍为多年生中生草本植物(占28.97%),说明在乡村地区的入侵植物更倾向于多年生中生型草本植物,这些植物可能具有较强的竞争能力和适应性,从而占据了较多的生境. 但是,城市河段的情况有所不同. 本土植物倾向于多年生中生草本植物,占总生活型的34.56%. 入侵植物倾向于一年生湿生草本植物(占31.21%),表明城市地区的本土和入侵植物在生活型上与乡村地区的本土和入侵植物相似,都是具有较长生命周期的中生型草本植物. 在城市河段中外来植物主要以一年生中生草本植物为主(占29.45%),表明城市环境对于一年生中生型草本植物的生存和繁殖更为有利,可能是由于城市的频繁扰动和人为活动,导致了这类植物更易于快速繁殖和适应环境. 通过对不同河段植物群落生活型构造的描述,可以了解到乡村地区和城市地区植物群落的生活型组成存在显著差异. 这种差异可能受到环境条件、人为干扰和竞争方式等因素的影响,进一步说明了不同区域植物群落的适应能力和生态特征各不相同.
-
由表 2可知,乡村和城市河段的乡土植物群落具有最高的相似度,为0.823,意味着乡村和城市河段的乡土植物之间有较高的物种共享性和相似性. 乡土植物的相似度高可能与它们存在共同的生态环境和种群适应性有关. 在乡村和城市河段中外来植物的相似度稍低,为0.692,表明在不同区域中外来植物的种群组成存在差异,可能受到不同环境条件和人为干扰的影响. 外来植物的相似度较高,说明其在乡村和城市环境下的适应能力相对较强. 本文研究结果进一步揭示了随着人类干扰活动增多,植物种类替换引发了群落组成的大幅度变化,从而降低了不同区域植物种类的相似度,表明人类活动对植物群落的影响显著,包括生境破坏、引入外来物种和改变生态过程等. 这些干扰因素导致群落物种组成变动,进而降低了植物种类的相似度. 根据相似度的高低排序,可以看出乡土植物在乡村和城市河段的相似性最高,其次是入侵植物,最低是外来植物. 这进一步验证了在不同区域中,植物群落的组成存在较大差异. 乡土植物和入侵植物在乡村和城市河段的相似度较高,可能是由于它们对当地环境的适应性较强所致. 外来植物种群的相似度较低,说明它们在不同区域的适应能力较弱,更容易受到人类活动的影响而发生变化.
本文利用Sorensen相似性指数对不同区域河段的植物群落构成进行对比分析,研究结果揭示了不同区域之间植物种类相似度存在差异. 随着人类活动干扰强度增加,导致了植物种类的替换和群落组成的变化,并降低了不同区域植物种类的相似度.
-
根据表 3的结果可知,不同区域段湿地外来入侵植物物种存在差异. 其中,各河段独有的外来植物数量相差不大,意味着不同湿地区域之间在外来植物入侵物种的种类数上没有明显的差异. 然而,尽管数量上相近,但具体的外来植物组成存在差异. 随着人类干扰活动增加,不同区域段的湿地植物组成发生了明显的变化. 特别是在城市段,国外外来物种数量比乡村段增长了大约2倍,表明城市段湿地更加容易引入国外外来物种,可能因为其受到了更多的人类活动干扰和外来物种传播途径的影响. 具体而言,城市段湿地中新出现的国外外来物种包括鸢尾、凤尾兰、绣球和夹竹桃,这些物种的引入与城市环境的特点紧密相关. 鸢尾和凤尾兰是受欢迎的观赏植物,很可能通过园艺市场进入城市段的湿地. 绣球和夹竹桃等物种可能通过人类带入的种子或苗木传播. 在城市环境下,人类活动和交通流动的频繁性为这些植物的传播提供了更多的机会. 相比之下,乡村段湿地的外来物种数量相对较少,这可能是由于乡村段受到的人类活动干扰较少,包括园艺、贸易和旅游等活动的影响较小所致. 因此,乡村段湿地的物种组成变化较小,外来物种数量较少. 城市段由于频繁的人类活动和交通流动,以及外来物种的传播途径更多,导致国外外来物种数量显著增长;而乡村段受人类活动干扰较少,外来物种的数量相对较少. 对于湿地生态系统的管理和保护,需要针对不同区域段的特点制定相应的措施.
3.1. 不同区域段湿地植物物种的构成及来源
3.2. 不同区域段湿地植物物种比例和优势度
3.2.1. 外来物种比例
3.2.2. 优势度与优势种
3.2.3. 生活型结构
3.3. 不同区域段湿地植物物种相似性分析
3.4. 不同区域段湿地外来入侵植物物种
-
人类活动对河岸湿地植物群落的影响是一个复杂的研究领域,涉及不同区域和植物物种来源的差异[11-12]. 本研究发现乡村段河岸上的植物以乡土植物为主,可能是因为乡村段河岸受到农业活动的较大影响[13]所致. 农业活动包括种植作物、养殖畜禽等,这些活动会导致土壤变化、水质污染和生物多样性减少. 乡村地区的农业活动通常以传统农业为主,种植的作物多为乡土植物,如稻谷、玉米等,这些植物适应当地气候和土壤条件,并且生长状况较好,因此在乡村段河岸上的植物群落中占比较高[14]. 同时,乡村段河岸上的植物群落受到人类活动的保护和管理[15]. 另外,乡村地区通常人口密度较低,人类对河岸湿地的干扰相对较小,乡村居民对当地自然环境有较高的认识和保护意识,通常采取措施来保护河岸湿地的植物群落,如禁止乱砍滥伐、限制过度放牧等,这些保护和管理措施有助于维持乡村段河岸上乡土植物的较高比例[16]. 乡村段河岸上的植物群落还受到土地利用方式的影响,乡村地区的土地利用方式主要以农业为主,农田通常占据河岸带的一部分,种植的作物多为乡土植物. 而城市地区的土地利用方式主要以工业和居住为主,河岸带通常被开发为城市公园或居民区,种植的植物多为观赏植物和外来植物.
本研究发现,乡村段河岸上的外来植物优势度显著低于城市段,这可能是因为乡村段的土地利用方式更多样化,包括农田、草地和林地,而城市段主要是城市建设用地所致[17]. 城市建设用地的人为干扰程度较高,土壤质量可能较差,限制了外来植物物种的生存和繁殖[18]. 相比之下,乡村段的土壤质量可能更适合乡土植物物种的生长,因此外来植物的优势度较低. 城市段的建设活动导致生境破坏和退化,如水体和土壤污染,这些环境污染对乡土植物物种的生长和繁殖造成了不利影响,但对适应能力较强的外来植物物种可能并不致命. 因此,在城市段的河岸上外来植物物种更容易适应并占据生境,形成优势[19]. 城市段的河岸受到更频繁和强烈的人为干扰,如园林绿化和道路建设,这些干扰可能导致乡土植物物种减少,而一些适应能力较强的外来植物物种却会受益于这些干扰[20]. 相比之下,乡村段的河岸受到人为干扰较少,乡土植物物种更容易维持其优势地位. 在城市环境中,由于人类活动的干扰和环境条件的改变,外来植物更容易适应和繁殖. 一年生中生草本植物具有快速生长和繁殖的特点,适应能力强,能够迅速占领空闲土地和受干扰的生境. 因此,在城市段中以一年生中生草本植物为主的外来植物比例较高. 这种现象可能是由于城市环境中土地利用变化、人为干扰和气候条件等多种因素所致[21-22]. 城市地区常常存在大量的空地和裸露土壤,为一年生中生草本植物提供了适宜的生长空间. 城市环境中的人为干扰,如建筑工地和道路施工会打乱土壤和植被的稳定性,进一步提供了一年生中生草本植物生长的机会[23].
本研究还发现乡村段与城市段本土植物的相似性最高,其乡村段与城市段的外来植物次之,这可能是由于地理距离和环境相似所致. 乡村段与城市段通常地理位置相近,意味着它们可能共享相似的自然环境,如土壤类型、气候特征和水源等,这些相似的环境条件为本土植物提供了类似的生存和繁殖条件,因此乡村段与城市段的本土植物种类相似性较高. 还可能是由于人类活动和人为引入所致,城市作为人类活动集中的地区,通常会引入许多外来植物种类,这些外来植物可能通过人类活动(如农业、园艺、贸易等)或自然传播进入城市段. 乡村段与城市段地理位置相近,可能存在相似的人类活动和自然传播途径,因此乡村段与城市段的外来植物种类也可能相似. 城市段频繁的人类活动,更有可能引入更多的外来植物种类.
随着人类活动增加,对自然环境的干扰不断加大. 这种干扰包括土地利用变化、森林砍伐、生物入侵和污染物排放等,对植物群落的物种组成和结构产生了深远的影响[24]. 这种影响导致了物种的更替变化,进而使不同区域的植物群落构成呈现出差异,降低了区域之间植物物种的相似性. 生物入侵是植物群落的重要干扰因素之一[25]. 通过外来物种侵入到原有生态系统中,并对原有物种和生态系统造成负面影响. 这些外来物种通常具有较强的适应能力和繁殖能力,容易在新环境中形成大规模的种群[26],它们与原有物种竞争资源,导致原有物种减少或灭绝. 在生物入侵过程中,植物群落的物种组成和结构发生了较大变化,新的物种取代了原有物种,导致植物群落的物种更替并产生差异[27]. 因此,生物入侵也是导致区域间植物物种相似性降低的重要原因,这种现象对生态系统的稳定性有着重要的影响,需要引起我们的重视和关注.
-
针对不同区域与不同植物物种来源条件下,人类活动对河岸湿地植物群落的影响研究表明,6个处理中乡村段河岸上乡土植物的比例最高(65.38%). 乡村段的乡土植物比城市段多,乡村段的入侵植物明显低于城市段. 无论乡村段还是城市段,乡土植物的优势度都最高,乡村段河岸的乡土植物的优势度高于城市段,而乡村段河岸外来植物的优势度明显低于城市段,乡村段河岸的入侵植物的优势度也显著高于城市段. 在乡村段的外来植物中,多年生旱生草本植物的比例最高(38.29%),而城市段的外来植物中一年生中生草本植物的比例最高(29.45%). 乡村段与城市段本土植物相似度最高(0.823),其次是乡村段与城市段的外来植物(0.692). 随着人类活动干扰强度增加,物种的更替变化导致不同区域植物群落构成呈现出很大的差异,降低了区域之间植物物种的相似性. 城市段出现了新的国外外来种,数量比乡村段增长了大约两倍,新出现的物种包括鸢尾、凤尾兰、绣球、夹竹桃等. 河南省唐河县段河岸带的外来植物种类构成特点,反映出人类活动对河岸带外来植物干扰逐渐扩大的趋势,包括快速城市化和农业活动等.
 DownLoad:
DownLoad: