


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
菜地由于复种指数高、施肥量大、灌溉频繁[1], 是大气CO2、CH4、N2O等温室气体的重要排放来源之一。我国作为目前世界上最大的蔬菜生产国, 蔬菜种植面积2 198.6万hm2, 占全国农作物总播种面积的13.03%[2]。据估算, 我国1 hm2菜地一年的温室气体排放量约为6 244 kg二氧化碳当量, 排放总量为13 728.06万t二氧化碳当量, 菜地温室气体排放量已经超过了我国谷物作物系统温室气体的排放量[3]。因此, 在全球气候变暖的背景下, 如何实现菜地在蔬菜增产的同时减少温室气体排放对于农业绿色发展具有重要意义。
菜地温室气体排放受这些气体在土壤剖面产生、消耗和传输作用的影响, 并受环境和管理因素的调控。Button等[4]通过不同土层间的CO2浓度梯度计算得来的小麦田理论排放通量仅与实际值相差6%;冯迪等[5]发现, 菜地N2O排放通量与土壤N2O含量呈显著的正相关关系;张凯莉等[6]发现, 菜地地表CH4浓度与10 cm、20 cm土层的CH4含量之间存在显著的正相关关系。通过观测菜地土壤剖面的温室气体含量, 可以深入理解温室气体在土壤剖面的分布特征, 有助于菜地温室气体减排研究。
地膜覆盖作为一种常用的农田管理措施, 可以有效改善土壤水热状况及理化性质, 影响土壤微生物群落的分布[7], 增加粮食产量[8-9]。有研究发现, 覆膜会影响土壤温室气体的产生, 在辣椒—萝卜轮作菜地, 覆膜显著提高了观测期间的土壤CO2含量以及萝卜季的N2O含量, 但显著降低了萝卜季土壤剖面的CH4含量[10]。生物炭C/N高, 比表面积大, 孔隙发达, 表面官能团丰富, 稳定性好, 可以改善退化和低肥力的土壤[11], 提高作物产量[12]。研究发现, 生物炭施加也会影响土壤温室气体的产生, 在小麦—水稻轮作农田, 施氮条件下配施生物炭降低了水稻季土壤上层剖面的CH4含量, 以及小麦季土壤剖面各层次和水稻季土壤上层剖面的N2O含量[13]。目前, 关于地膜覆盖和生物炭施加对菜地土壤剖面温室气体含量的影响研究还鲜见报道。基于此, 本研究以西南地区典型的辣椒—萝卜轮作菜地为研究对象, 探究地膜覆盖、生物炭施加及两者的交互作用对菜地土壤剖面温室气体体积分数的影响, 以期为西南地区菜地温室气体减排提供参考。
全文HTML
-
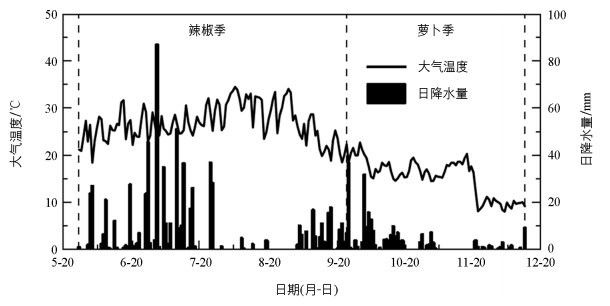
试验菜地位于西南大学实验农场(30°26′ N, 106°26′ E), 海拔230 m, 属于典型的亚热带湿润季风气候。试验期间(2020年5月-2020年12月)的气温和降水量如图 1所示。试验区整体上雨热同期, 气温、降水量具有明显的季节变化规律, 辣椒季的气温和降水量高于萝卜季。辣椒季的平均气温为26.90 ℃, 累积降水量为718.06 mm;萝卜季的平均气温为15.35 ℃, 累积降水量为224.79 mm。试验菜地土壤为中生代侏罗系沙溪庙组灰棕紫色沙泥岩母质上发育的中性紫色土, 土壤表层(0~20 cm)的理化性质为:土壤pH值6.83、有机质25.29 g/kg、总磷646.00 mg/kg、有效磷5.94 mg/kg、总钾9.44 g/kg、有效钾102.47 mg/kg、全氮1.35 g/kg、碱解氮56.62 mg/kg。
-
试验种植模式为西南地区常见的辣椒—萝卜轮作, 辣椒品种为‘龙椒一号’, 购于重庆龙胜种子经营部, 萝卜品种为‘长白玉’, 购于绵阳市涪城区龙门镇农资批发市场。本试验采用的生物炭由玉米秸秆制成, 购于南京三聚生物质新材料科技有限公司, 其基本理化性质为:比表面积224.25 m2/g、有机碳537.97 g/kg、总磷2.00 g/kg、总钾27.15 g/kg、全氮0.61 g/kg。生物炭于试验开始前一次性施入, 之后不再添加。试验开始前, 先将土地深翻, 使土地尽可能平整, 然后平地起垄, 垄长2.8 m, 垄宽0.8 m, 垄高0.15 m, 垄间距0.4 m。地膜选用0.005 mm厚的聚乙烯透明塑料薄膜平整覆于垄上, 两边用细土压实。试验在不覆膜(NF)和覆膜(F)处理下各设置了3种不同的生物炭施加水平(B0:0 t/hm2;B20:20 t/hm2;B40:40 t/hm2), 共6个处理, 分别为NFB0、NFB20、NFB40、FB0、FB20、FB40。每个处理3个重复, 共18个小区。
因不可抗因素, 辣椒错过最佳播种时间, 于2020年5月27日移栽辣椒苗, 6月21日追肥, 之后辣椒成熟分批收获, 并于9月2日收获最后一批。萝卜籽播种于2020年9月24日, 10月23日追肥, 12月24日萝卜成熟后一次性收获。所有小区在辣椒季和萝卜季的施肥量相同, 氮肥(尿素)、钾肥(氯化钾)、磷肥(过磷酸钾)施加量分别为300、100、150 kg/hm2, 磷肥和钾肥作为基肥在播种或移栽前一次性施加, 氮肥在基肥和追肥时各施加一半。试验期间田间管理方式与当地种植习惯相同, 蔬菜种植期间无人工灌溉。
-
采用扩散箱法采集不同处理下菜地土壤剖面的温室气体。扩散箱由集气管、导气管、橡胶塞、三通阀4个部分组成。集气管是一根圆柱形的PVC管(直径5 cm, 容积1 000 cm3), 管壁一侧钻有孔洞(4排, 每排25个, 直径3 mm), 管端两侧用橡胶塞密封, 确保集气管只能通过孔洞交换气体;导气管是一根内径为1 mm的聚乙烯塑料管, 其一端穿过橡胶塞伸到集气管中央, 另一端垂直伸出地表并在末端配有三通阀, 非采集时间三通阀保持关闭状态, 防止集气管内气体通过导气管泄露。试验开始前, 将扩散箱洞口向下水平埋在10、20、30 cm土深处, 确保集气管中心轴距垄上表面的距离即为预埋深度, 以减小试验误差。
试验观测期间, 每月选择一晴朗天气在上午的9:00-11:00进行气体样品的采集。研究表明, 这个时期的土壤温室气体通量能够代表一天的平均值, 因为这个时期的土壤温度接近土壤温度的日均值[14]。由于土壤温室气体通量受温室气体在土壤剖面产生、消耗和传输作用的影响, 本研究土壤温室气体体积分数的观测也选在这一时间段内进行。采样时, 用60 mL注射器从扩散箱采集不同剖面的气体, 采集气样后关闭三通阀, 并在24 h内将气样送至实验室使用安捷伦气相色谱仪(Agilent 7890A)测定CO2、CH4、N2O体积分数。CO2和CH4体积分数使用氢火焰离子检测器(FID)测定, N2O体积分数使用电子捕获检测器(ECD)测定, 具体分析方法及工作条件参照文献[15]。
-
每月采集一次垄上土壤样品, 采样深度为0~20 cm。采样时, 使用“四分法”将每个采样点采集的5份样品均匀混合为1个土样, 剔除土样中的杂物, 装入自封袋。一部分进行风干处理, 另一部分放入4 ℃冰箱内保存。土壤有机碳(SOC)采用重铬酸钾氧化—外加热法测定, 全氮(TN)采用凯氏定氮法测定, 可溶性有机碳(DOC)采用Multi N/C 2100分析仪测定, 硝态氮(NO3--N)和铵态氮(NH4+-N)经2M KCl溶液浸提后用流动注射分析仪(FIAstar 5000, 德国)测定。土壤样品在105 ℃的烘箱中干燥48 h后计算土壤质量含水率。在每次采集土样的同时, 使用JM624型便携式温度计测量土壤地下5 cm温度。
-
采用IBM SPSS Statistics 26进行方差分析。同一生长季, 不同处理下不同土层间温室气体体积分数的差异显著性采用三因素方差分析;同一生长季, 不同处理下各环境因子的差异显著性采用双因素方差分析。p<0.05视为差异性显著。
1.1. 试验区概况
1.2. 试验设计与方法
1.3. 气体样品采集与测定
1.4. 土壤环境因子的测定方法
1.5. 数据处理
-
观测期间不同处理下的地下5 cm温度及土壤质量含水率如表 1所示, 地下5 cm温度表现为辣椒季高于萝卜季, 这与气温的季节分布一致, 土壤质量含水率则表现为辣椒季低于萝卜季。无论是在辣椒季, 还是在萝卜季, 地膜覆盖、生物炭施加及两者的交互作用对地下5 cm温度和土壤质量含水率均无显著影响(p>0.05)。
-
观测期间不同处理下的土壤NH4+-N、NO3--N及TN质量分数如表 2所示。在辣椒季, 无论是NH4+-N, 还是NO3--N, 其在不同处理下的变化规律均表现为:NFB0<FB0, NFB20<FB20, NFB40<FB40。覆膜显著提高了NH4+-N(p<0.05)和NO3--N(p<0.01)质量分数, 但生物炭施加及其与覆膜交互作用的影响则不显著(p>0.05)。不同处理下的TN质量分数表现为:NFB0<NFB40<NFB20, FB0<FB20<FB40。无论覆膜与否, 生物炭施加均显著提高了TN质量分数(p<0.05), 但覆膜及其与生物炭施加交互作用的影响不显著(p>0.05)。在萝卜季, 覆膜、生物炭施加及两者的交互作用对NH4+-N、NO3--N以及TN质量分数均无显著影响(p>0.05)。
-
表 3显示了不同处理下土壤SOC和DOC质量分数。不论是在辣椒季, 还是在萝卜季, 不同处理下的SOC和DOC质量分数均表现为:NFB0<NFB20<NFB40, FB0<FB20<FB40。无论覆膜与否, 随着生物炭施入量的增加, 土壤SOC质量分数(p<0.01)和DOC质量分数(p<0.05)均显著增加。然而覆膜及其与生物炭施加的交互作用对SOC和DOC质量分数却均无显著影响(p>0.05)。
-
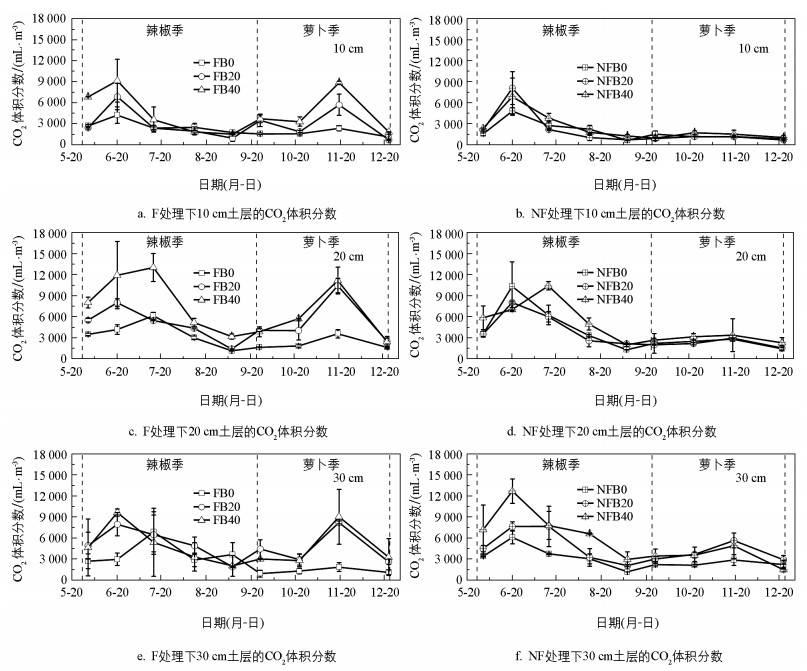
在不同处理下, 10、20、30 cm土层的CO2体积分数具有相似的季节变化规律, 整个观测期间的CO2体积分数为653.55~13 059.21 mL/m3(图 2)。辣椒季, CO2体积分数先升高后降低, 峰值大多出现在6月中旬。萝卜季, CO2体积分数也呈先升高后降低趋势, 峰值均出现在11月中旬。
表 4显示了不同处理下菜地土壤剖面的CO2体积分数。在萝卜季, 相比于NF, F显著提高了CO2体积分数(p<0.01), 增幅为60.62%, 而辣椒季覆膜的影响则不显著(p>0.05)。不同生物炭施加量下的CO2体积分数在辣椒季和萝卜季均表现为:B0<B20<B40。其中, 相比于B0处理, B20和B40处理在辣椒季使CO2增加了21.08%和64.60%, 在萝卜季增加了80.42%和103.63%。施加生物炭显著提高了辣椒季和萝卜季的CO2体积分数(p<0.01)。无论是辣椒季还是萝卜季, 土壤剖面CO2均随土壤深度的增加而显著增加(p<0.01)。相比于10 cm土层, 20 cm和30 cm土层的CO2在辣椒季分别增加了72.50%和60.19%, 在萝卜季分别增加了63.72%和58.35%。另外, 三因素方差分析结果表明, 除覆膜与生物炭施加、覆膜与土壤深度的交互作用对萝卜季的CO2有显著影响(p<0.01)外, 覆膜、生物炭施加以及土壤深度的交互作用对整个观测季的CO2均无显著影响(p>0.05)。
-
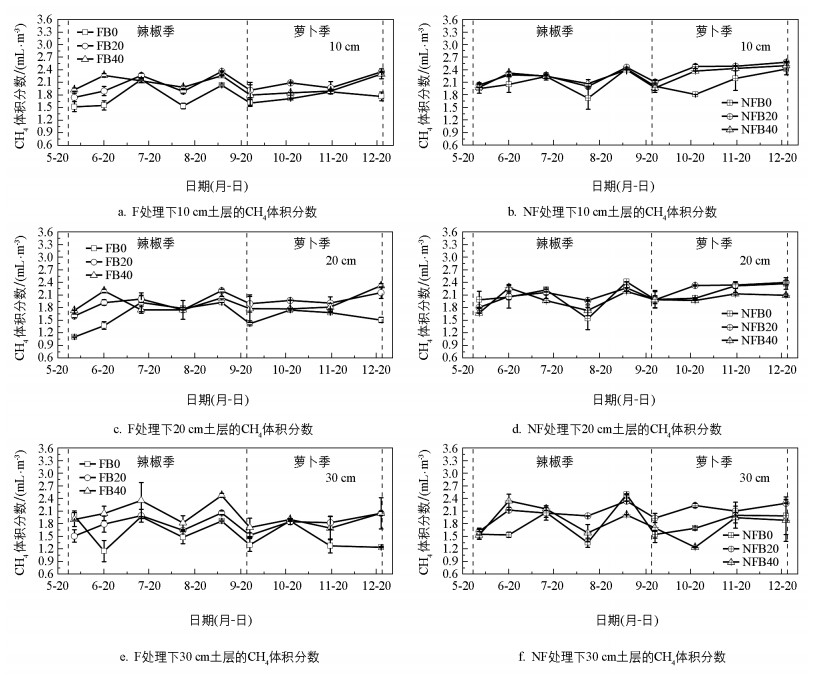
覆膜与生物炭施加均没有改变土壤CH4体积分数随季节的变化趋势(图 3), 整个观测期间的CH4体积分数为1.10~2.59 mL/m3。辣椒季, 各土层CH4体积分数随着时间推移波动变化, 大致在6月中旬和9月上旬出现两个峰值;萝卜季, 各土层的CH4体积分数随着时间推移呈轻微上升趋势。
表 5显示了不同处理下菜地土壤剖面的CH4体积分数。无论是辣椒季还是萝卜季, 覆膜、生物炭施加、土壤深度均对CH4体积分数产生了显著影响。相比于NF, F显著降低了辣椒季(p<0.05)和萝卜季(p<0.01)的CH4体积分数, 降幅分别为7.84%和13.74%;相比于B0, 生物炭施加显著提高了整个观测期间的CH4体积分数, B20、B40处理下的CH4体积分数在辣椒季分别提高了9.24%和10.87%(p<0.05), 在萝卜季分别提高了17.13%和8.84%(p<0.01);无论是辣椒季还是萝卜季, 不同土层间的CH4体积分数从小到大均为:30 cm(1.90 mL/m3和1.79 mL/m3)、20 cm(1.92 mL/m3和2.00 mL/m3)、10 cm(2.07 mL/m3和2.11 mL/m3), 随着土壤深度的增加, 辣椒季(p<0.05)和萝卜季(p<0.01)的CH4体积分数均显著降低。另外, 三因素方差分析结果表明, 除覆膜与生物炭施加的交互作用对萝卜季的CH4有显著影响(p<0.05)外, 覆膜、生物炭施加以及土壤深度的交互作用对整个观测季的CH4均无显著影响(p>0.05)。
-
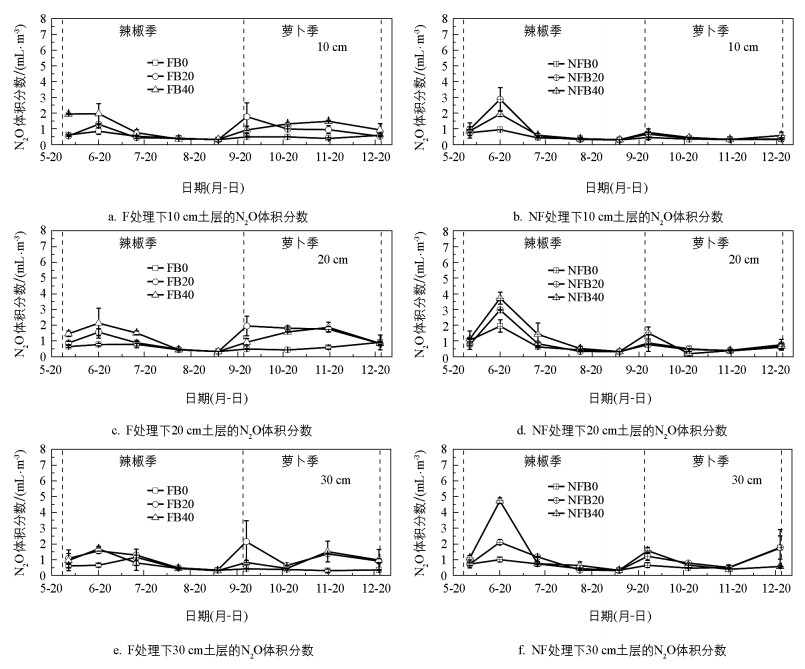
在不同处理下, 10、20、30 cm土层的N2O体积分数具有相似的季节变化规律, 整个观测期间的N2O体积分数为0.20~4.76 mL/m3(图 4)。辣椒季, N2O体积分数先升高后降低, 峰值均出现在6月中旬;萝卜季, N2O体积分数总体变化平稳, 无太大波动。
表 6显示了不同处理下菜地土壤剖面的N2O体积分数。在萝卜季, 相比于NF, F显著提高了N2O体积分数(p<0.01), 增幅为47.76%, 而辣椒季覆膜的影响不显著(p>0.05)。无论是辣椒季还是萝卜季, B20和B40处理下的N2O体积分数相比于B0处理均显著提高。其中, B20和B40处理在辣椒季使N2O体积分数增加了37.88%和75.76%(p<0.05), 在萝卜季增加了77.19%和61.40%(p<0.01)。不同于CO2和CH4, 整个观测期间N2O体积分数随土壤深度增加而不显著地增加(p>0.05)。三因素方差分析结果还表明, 除覆膜与生物炭施加、覆膜与土壤深度的交互作用对萝卜季的N2O影响极显著(p<0.01)外, 覆膜、生物炭施加以及土壤深度的交互作用对N2O均无显著影响(p>0.05)。
2.1. 土壤环境因子
2.1.1. 土壤地下5 cm温度和质量含水率
2.1.2. 土壤氮质量分数
2.1.3. 土壤碳质量分数
2.2. 土壤剖面CO2体积分数
2.3. 土壤剖面CH4体积分数
2.4. 土壤剖面N2O体积分数
-
本研究发现, 无论是在辣椒季还是在萝卜季, 所有处理均表现为CO2体积分数随土壤深度增加而增加(表 4)。这主要是因为, 虽然表层土壤有机质含量丰富, 有利于CO2的生成, 但丰富的有机质改变了土壤结构, 增加了土壤的渗透性[16], 使得表层土壤的CO2更易于向大气扩散。而深层土壤结构更加紧密, 气体有效扩散系数相对较低[4], CO2向周边土层的扩散受到限制。有田间试验发现, 在华北平原冬小麦—夏玉米农田, 土壤CO2含量随深度的增加而增加[17], 与本研究结果相似。
在萝卜季, 覆膜提高了土壤CO2体积分数(表 4), 这可能是由于地膜覆盖可以物理阻隔CO2向大气扩散。但在辣椒季, 覆膜对土壤CO2却无显著影响(表 4), 与萝卜季的结果不一致。Nishimura等[18]研究发现, 一方面土壤温室气体可以从覆膜的垄间向相邻未覆膜的沟水平扩散, 另一方面随着环境温度增加, 从地膜垂直渗出的温室气体量呈指数增加。在本试验中, 辣椒种植于夏秋季, 天气炎热, 各处理下的土壤平均温度为25.34 ℃, 远高于萝卜季的14.58 ℃(表 1), 这使得辣椒季的CO2水平扩散速率以及垂直渗出速率要大大高于萝卜季, 可能就是这种温度差异导致辣椒季和萝卜季在研究结果上存在差异。
在辣椒季和萝卜季, 生物炭施加均显著提高了土壤CO2体积分数(表 4)。这可能主要是有以下几个原因:第一, 生物炭促进了作物生长, 增强了作物吸收大气CO2的能力, 使植物根系向土壤中释放的碳增多[19]。本研究发现, 在辣椒季和萝卜季, 生物炭施加提高了SOC质量分数(表 3)。第二, 有研究发现, 当土壤施加生物炭后, 会促使其中的不稳定碳释放出来[20], 本研究的试验结果也表明, 在辣椒季和萝卜季, 生物炭施加提高了DOC质量分数(表 3)。SOC和DOC为微生物生长、繁殖提供了碳源, 促进了土壤呼吸。第三, 生物炭的多孔结构及其表面官能团的存在使得生物炭可以吸附土壤中的CO2[21-22], 防止CO2向大气扩散。第四, Kuzyakov等[23]研究发现, 新鲜生物炭虽然主要由芳香族化合物组成, 但也含有一定量的多糖及脂质, 使得生物炭本身可作为碳源而被缓慢地分解, 有利于CO2的生成。
-
与CO2相反, CH4体积分数随土壤深度的增加而降低(表 5), 这说明菜地是大气CH4的吸收汇[24]。当菜地表层吸收大气CH4后, 向下扩散, 由于底层土壤越来越紧实, 从而导致底层CH4体积分数低于表层。Chen等[10]通过田间试验也发现了与本研究相似的试验现象, 即较深土壤中的CH4体积分数显著低于表层土壤。然而, 杨波等[25]通过在稻麦轮作农田开展田间试验发现, 小麦生长季的CH4体积分数随着土壤深度增加而增加, 与本研究结论不一致。这可能是因为小麦季之前是水稻季, 由于水稻季往复淹水, 阻止了大气—土壤界面间O2的交换, 导致农田土壤可能形成产CH4所需的厌氧环境。到了小麦季, 虽然大气中的O2可以进入表层土壤, 但由于土壤底层气体有效扩散系数相对较低[4], 导致底层土壤可能仍是厌氧环境, 有利于CH4产生。
本研究发现, 覆膜显著降低了土壤CH4体积分数(表 5)。有田间试验发现, 在黄土高原玉米农田, 覆膜显著降低了土壤不同深度处的CH4体积分数[26], 与本研究结论相似。一方面, 地膜覆盖的阻隔作用限制了大气中的CH4进入土壤;另一方面, 地膜覆盖有效阻止了雨水对菜地的直接冲刷作用, 提高了蔬菜地NH4+-N和NO3--N质量分数(表 2), 可能造成了以下3个影响:第一, NH4+-N质量分数的增加可以促进CH4氧化[27], 进而降低土壤CH4浓度;第二, NO3--N为反硝化过程提供了底物, 有利于反硝化细菌增加, 这可能会与产甲烷菌形成微生物竞争[28];第三, 覆膜阻止了O2在土壤—大气界面中的交换, 使得菜地土壤可能形成局部厌氧环境。在厌氧环境下, 甲烷氧化菌利用NO3--N作为电子供体, 发生反硝化型甲烷厌氧氧化反应[29]。
本研究还发现, 生物炭的施加显著提高了土壤CH4体积分数(表 5)。这可能主要是由以下几个原因所致。首先, 生物炭孔隙率高, 施加土壤后可以增加土壤的通气性, 降低土壤的堆积密度[30], 有利于大气CH4进入土壤, 且生物炭可以吸附土壤中的CH4, 防止CH4从土壤中逸出。其次, 生物炭施入土壤后有利于大团聚体的形成[31]。由于大团聚体内的孔隙大小不一, 部分孔隙在大团聚体形成过程中被封闭, 外部空气与大团聚体的交流受到孔隙网络的限制, 缺氧成为团聚体最显著的环境特征, 这抑制了大团聚体内甲烷氧化菌的活性[32], 从而削弱了CH4的氧化作用。最后, 生物炭施加提高了菜地SOC和DOC的质量分数(表 3), 在厌氧或者局部厌氧环境下有利于CH4的产生。
-
与CO2相似, N2O体积分数随土壤深度的增加而增加(表 6)。贾俊香等[33]通过田间大棚试验发现, 菜地土壤剖面的N2O体积分数呈现50 cm>30 cm>15 cm>7 cm的梯度特征, 与本研究结论相似。造成这种现象的主要原因是:一方面, 与表层相比, 底层土壤更加紧密[4], 这阻止了底层N2O向周边土层扩散, 使得N2O易于在底层积累。另一方面, 硝化作用是菜地N2O产生的主要来源, 土壤N2O体积分数与土壤含水率呈显著正相关[34]。杜康等[35]通过研究发现, 在0~50 cm土层, 农田土壤含水量随着深度的增加而增加, 与N2O体积分数的分布趋势一致。
覆膜显著提高了萝卜季的N2O体积分数, 而对辣椒季的N2O体积分数无显著影响(表 6), 这与覆膜对土壤CO2的影响一致(表 4)。不同于菜地土壤吸附大气CH4, N2O同CO2一样, 都表现为从土壤向大气扩散的特征。因此, 覆膜对萝卜季和辣椒季N2O体积分数的影响不一致, 也如前文在CO2部分阐释的原因相同, 即:一方面, 覆膜有效阻止了土壤N2O向大气扩散。另一方面, 萝卜季和辣椒季土壤温度的差异导致N2O水平扩散量及垂直渗出量存在差异。
本研究发现, 无论辣椒季还是萝卜季, 生物炭施加均提高了土壤N2O体积分数(表 6)。王晓辉等[36]于温室中研究发现, 当生物炭施入菜地土壤后, 首先提高了土壤pH值, 增加了氨氧化古菌和氨氧化细菌群落的丰度, 增强了土壤的硝化潜势。且本研究中生物炭施加增加了菜地SOC和DOC的质量分数(表 3), 为氨氧化古菌、氨氧化细菌和反硝化细菌生长、繁殖提供了所需的碳源, 也为硝化过程提供了足够多的电子供体, 从而促使更多的铵态氮和硝态氮转为N2O。其次, 生物炭施入提高了nirK型反硝化细菌的基因丰度[36], 有利于反硝化过程中N2O的积累。最后, 同CO2和CH4一样, 生物炭的多孔结构有利于吸附土壤剖面产生的N2O, 防止产生的N2O从土壤中逸出。然而, 马芸芸等[37]发现, 秸秆生物炭施入水稻田后降低了剖面N2O体积分数, 与本研究结论不一致。这可能是因为水稻田长期处于淹水环境而导致土壤缺氧, 从而有利于反硝化反应。有研究发现, 在较好的反硝化条件下, 生物炭施入使得N2O/(N2O+N2)比值持续降低, 促使更多的N2O还原为N2[38]。
-
(1) 土壤剖面的CO2体积分数随土壤深度的增加而增加;覆膜提高了萝卜季土壤CO2体积分数, 但对辣椒季土壤CO2体积分数无显著影响;施加生物炭提高了整个观测期间的土壤CO2体积分数。
(2) 土壤剖面的CH4体积分数随土壤深度的增加而降低;覆膜降低了土壤CH4体积分数, 而施加生物炭则提高了土壤CH4体积分数。
(3) 土壤剖面的N2O体积分数随土壤深度变化不大;覆膜提高了萝卜季土壤N2O体积分数, 但对辣椒季土壤N2O体积分数无显著影响;施加生物炭提高了整个观测期间的土壤N2O体积分数。