



 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
在自然界的物种中,基因突变会产生某一类突变体植株,这类植株的性状主要表现在无明显病原菌侵染、机械损伤等相关条件刺激下,茎秆、叶片、叶鞘、穗等部位会自发产生坏死斑点,其表型特征与过敏反应(Hypersensitive Response,HR)相似,但不存在致病因子,人们将这类突变体统称为类病斑突变体(Lesion Mimic Mutants,LMMs)[1-2]。一般植物类病斑突变体内会引起大量过氧化物(Reactive Oxygen Species,ROS)积累、相关防御基因表达上调、细胞程序性死亡(Programmed Cell Death,PCD)、植保素激增等自发防御反应[3-4]。
水稻在生长发育过程中会遭遇多种病害威胁,主要包括白叶枯病、稻瘟病、纹枯病、白粉病等,而在我国南方地区稻瘟病和白叶枯病为常见的两种水稻植株病害。其中,水稻稻瘟病是由稻瘟病菌(Magnaporthe oryzae)引起的一种真菌性病害,全球每年因该病造成的产量损失有10%~30%,严重时竟会导致稻谷颗粒无收[5-6]。白叶枯病则是由革兰氏阴性黄单胞杆菌致病变种(Xanthomonasoryzae pv. oryzae)引起的细菌性病害,也是我国南方稻区主要的细菌性病害,严重降低水稻的光合作用,对稻米产量及品质具有较大影响[7-9]。研究表明,类病斑突变体植株往往表现出对稻瘟病及白叶枯病抗性效果显著[10-12],如rbl1通过影响磷脂酰肌醇的合成来控制PCD与免疫反应,突变体植株对稻瘟病菌和白叶枯病菌的抗性明显优于野生型[13],spl33编码一个eEF1A-like蛋白,能够负调控植物的免疫反应,其功能缺失使spl33对病原稻瘟病和白叶枯病具有广谱抗性[14],lmm8存在原卟啉原氧化酶(Protoporphyrinogen Oxidase,PPOX)的基因缺陷而表现出对稻瘟病菌与白叶枯病菌的高抗性等[15]。学者们认为研究导致水稻类病斑发生的基因是破译防御途径的关键,有助于提高植物广谱病原体抗性。
本研究的前期工作已获得遗传性状稳定的类病斑突变体spl45,并对突变体spl45进行了一系列性状考察及相关实验,可为深入探究候选目的基因的分子调控机制,创制优良抗病水稻种质资源提供参考。
全文HTML
-
突变体植株spl45,利用甲基磺酸乙酯(Ethyl Methane Sulphonate,EMS)诱变剂处理籼型杂交水稻Xinong 1B获得,经世代自交后突变体植株表型稳定。将突变体spl45与Jinhui 10杂交所得F1代再自交获得F2代,用于基因定位。
-
于2023年4月在西南大学水稻研究所歇马基地进行材料种植,分别栽种野生型(WT)Xinong 1B及突变体spl45植株,采用常规水稻种植管理方式,从苗期开始观察突变体与野生型植株的表型差异,拍照记录类病斑出现的时期及形态特征。
-
在植株完全成熟后,分别随机选取10株WT、10株突变体spl45植株,对其株高、穗长、穗粒数、结实率等农艺性状进行详细调查,并对这些数据结果进行t检验分析。
-
选择分蘖期具有代表性的WT和突变体spl45植株,参照Arnon[16]和张晓艳等[17]的方法进行光合色素含量测定,剪碎剑叶、倒二叶、倒三叶后称取0.1 g放入10 mL丙酮和纯水混合液(V/V=4∶1)试管中,黑暗处理48 h期间多次振荡后,测定663 nm、645 nm、470 nm处的吸光度值,计算光合色素含量。
利用便携式光合作用测定仪(Li-6800)测定抽穗期剑叶、倒二叶、倒三叶叶片的光合参数。每组样品重复3次,野生型和突变体结果进行t检验分析。
-
植株分蘖期分别在WT和突变体spl45的剑叶中部剪取约1 cm长的叶片,石蜡包埋后用冷冻切片机将其切成25 μm薄片,将薄片放置在载玻片用ddH2O反复冲洗,使载玻片中仅有叶片薄片,盖上盖玻片,在荧光显微镜(OLYMPUS,18A16537)下观察拍照。
-
对分蘖期WT及突变体spl45的倒二叶进行遮光。以WT为对照,用宽度为2 cm的锡纸对spl45未产生斑点的叶片进行遮光处理,7 d后将锡纸摘除并拍照,复光7 d后再次拍照观察。
-
台盼蓝染色:在植株分蘖期剪取有表型的WT和突变体spl45叶片,然后将叶片浸泡在台盼蓝溶液中,真空70 ℃水浴锅加热2 min后冷却,在室温下放置12 h后,再用25 mg/mL水合氯醛溶液进行脱色。脱色后进行拍照记录。
二氨基联苯胺(3,3′-diaminobenzidine,DAB)染色:植株分蘖期剪取有表型的WT和突变体spl45叶片进行二氨基联苯胺(DAB)染色,将叶片放入1 mg/mL的DAB染液中,真空处理15 min后避光室温下放置20 h,然后将DAB染液倒掉,再将叶片浸泡在无水乙醇中进行水浴脱色,更换无水乙醇反复脱色使叶绿素完全去除,观察叶片中褐色斑点的相应位置,拍照保存。
脱氧核苷酸末端转移酶介导的dUTP缺口末端标记(TdT-mediatedd UTP Nick-END Labeling,TUNEL)染色:取分蘖期WT和突变体spl45的叶片放入甲醛-乙酸-乙醇(FAA)固定液中室温放置24 h,再放入装有包埋剂(Tissue-Tek,SAKURA)的离心管中,待包埋剂完全凝固,利用切片机(Thermo Scientific,USA)切至6~12 μm厚度蜡片,然后将蜡片移至载玻片上用ddH2O展开,再置于42 ℃烘箱3 d,除去多余水分。依次进行脱蜡、透膜、末端标记、DAPI染色,吸去载玻片液体,封片,利用激光共聚焦显微镜拍照,方法参照DeadEnd荧光TUNEL系统试剂盒(Promega,美国)执行。
-
在分蘖阶段随机选取3株突变体spl45和WT倒二叶,取0.2 g,使用南京建成生物公司的试剂盒来检测过氧化氢(hydrogen peroxide,H2O2)、过氧化氢酶(catalase,CAT)、过氧化物酶(peroxidase,POD)和超氧化物歧化酶(superoxide dismutase,SOD)的含量,结果进行t检验分析。
-
在西南大学水稻研究所歇马基地内对WT和突变体spl45进行稻瘟病抗谱测定及白叶枯病抗性鉴定检验。参照黄富等[18]的方法进行稻瘟病菌接种液制备,对3叶期水稻幼苗进行单孢悬浮液喷施接种稻瘟病,田间抗病等级评价参照国际水稻研究所(IRRI)水稻稻瘟病评级系统标准[19]。再参照孙富等[20]的方法对WT和突变体spl45进行水稻白叶枯病人工接种鉴定。
-
以Jinhui 10为母本、以突变体spl45为父本进行杂交获得F1代,而后F1自交获得F2代群体,利用卡方检测分析F2群体的分离比。构建基因混池,利用实验室已备差异引物进行连锁位点的初步确认,在初定位区间内设计indel标记进行基因精细定位,引物序列如表 1所示。
-
基因表达分析使用KKFast Plant RNAPURE Kit(庄盟生物)提取野生型和突变体spl45的RNA,用RT MIX with DNase Kit(yuyi-Landi生物,中国)进行逆转录。所得产物稀释20倍后,使用CFX Connect系统(Bio-Rad,美国)和SYBR Premix Ex Taq II试剂盒(TaKaRa,日本)进行反转录定量分析(Quantitative Reverse Transcription Polymerase Chain Reaction,qRT-PCR),设置3个生物重复。所有相关基因表达引物均参照Gramene数据库(
http://www.Gramene.org/ )中的基因序列,以OsActin1为内参基因进行设计,定量引物序列如表 2所示。采用GraphPad Prism 8.0.2(
http://www.graphpad.com/ )软件进行统计分析。并用t检验对实验数据进行验证。
1.1. 实验材料
1.2. 实验方法
1.2.1. 突变体spl45的表型分析
1.2.2. 农艺性状考察
1.2.3. 光合色素含量和光合参数测定
1.2.4. 冷冻切片
1.2.5. 遮光处理
1.2.6. 组织化学染色
1.2.7. 生理指标测定
1.2.8. 稻瘟病和白叶枯病抗病性鉴定
1.2.9. 突变体遗传分析与基因精细定位
1.2.10. 目的基因和病程相关基因的表达分析
-
WT和突变体spl45的表型鉴定如图 1a-d所示。突变体spl45在三叶期从最下部叶片开始出现褐色类病斑,由叶尖逐渐向内扩散至叶鞘,随着时间的推移,坏死斑点数目越来越多,至分蘖期除新生叶外,其余叶片及叶鞘均出现类病斑表型,并且突变体spl45植株还表现出明显的早衰和矮化现象。
-
对WT和突变体spl45的主要农艺性状数据,如株高、穗长、有效穗数、每穗粒数等进行统计分析(表 3),发现相较于WT,突变体spl45的株高、穗长、有效穗数、千粒质量和结实率都呈现出极显著下降状态(p<0.01),分别降低了16.7%、11.2%、19.9%、19.6%、50.0%,但在每穗粒数方面无统计学意义。
-
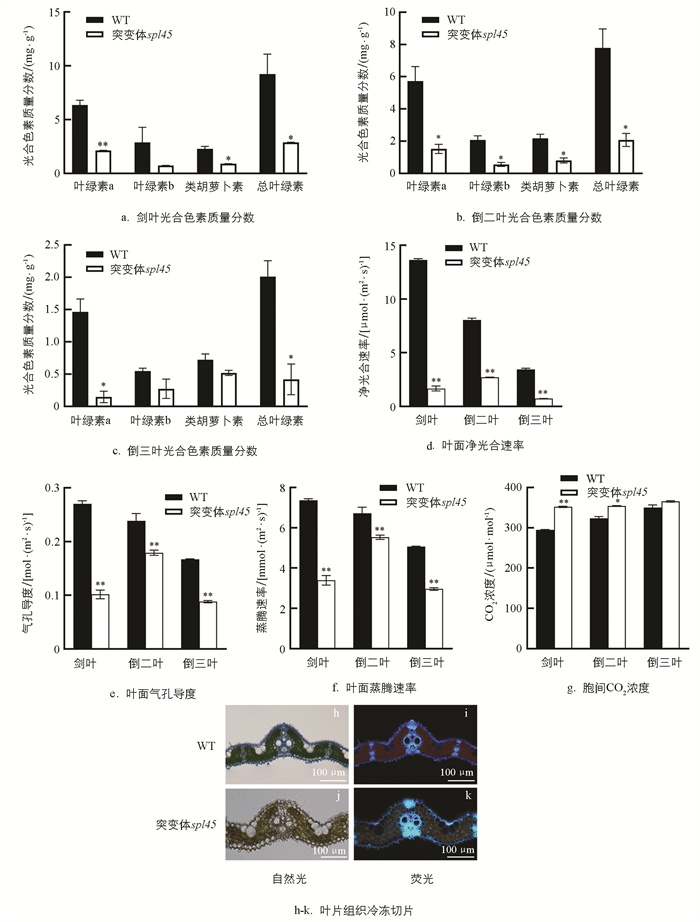
在WT和突变体spl45植株的抽穗期,对剑叶至倒三叶的光合色素质量分数进行检测。结果显示,突变体spl45植株剑叶、倒二叶、倒三叶中的叶绿素a、叶绿素b、类胡萝卜素和总叶绿素的光合色素质量分数都显著低于WT(图 2a-c)。与此同时,突变体spl45植株剑叶、倒二叶、倒三叶的净光合速率、气孔导度及叶面蒸腾速率效果也显著低于WT,而突变体spl45植株剑叶和倒二叶的胞间CO2浓度却明显升高(图 2d-g)。这些结果表明,与WT相比突变体spl45光合能力减弱。
为了进一步验证突变体spl45的光合能力相较于WT减弱,本研究对分蘖期的WT和突变体spl45叶片组织进行冷冻切片,发现突变体spl45在自然光下叶绿体明显失绿。在荧光显微镜下进行观察,发现其叶片中叶绿体自发产生的红色荧光强度明显弱于WT(图 2h-k),说明突变体spl45的叶绿体出现严重受损,光合能力减弱。
-
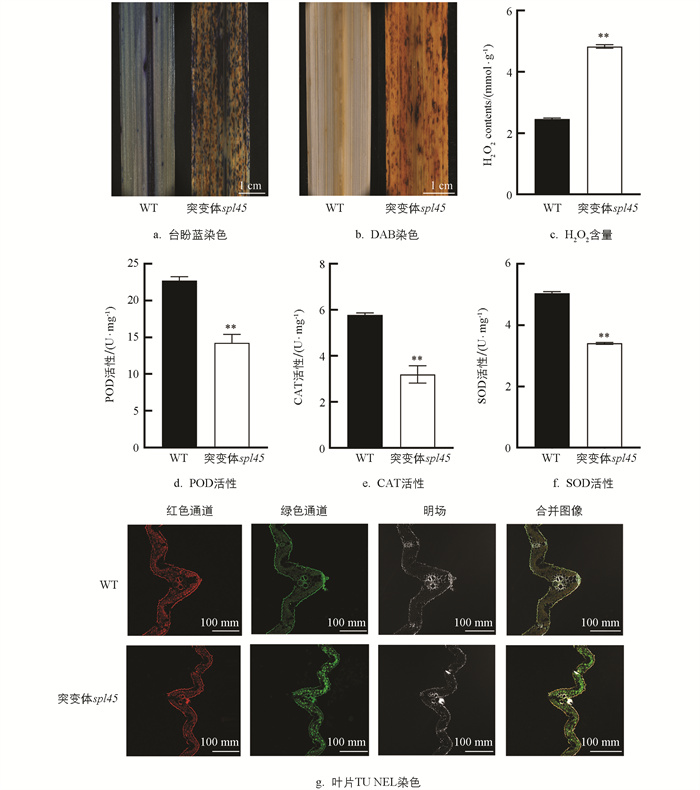
对分蘖期WT和突变体spl45进行了台盼蓝染色与DAB染色比较,以深入了解突变体spl45所引发的生理生化反应。台盼蓝染色结果显示,突变体spl45叶片的类病斑区域部位及周边被染成了深蓝色,而WT叶片则呈现出均匀的浅蓝色(图 3a),意味着突变体spl45叶片上出现了严重的细胞程序性死亡现象。通过对突变体spl45叶片的DAB染色,可以观察到病斑区域有严重的褐色斑点出现(图 3b),表明突变体spl45叶片上积累了大量的过氧化物;通过对WT和突变体spl45叶片进行H2O2含量和过氧清除酶活性测定,发现突变体spl45的H2O2含量显著高于WT(图 3c),与DAB染色结果一致;其POD、CAT和SOD的活性显著低于WT(图 3d-f)。由图 3可知,突变体spl45在清除活性氧方面的能力有所降低,进一步导致了大量过氧化物积累,从而触发了细胞的程序性死亡。
为进一步验证在突变体spl45出现的类病斑表型是细胞程序性死亡,本研究对材料叶片进行了TUNEL检测。结果显示,在WT叶片细胞中几乎没有绿色荧光信号存在,而突变体spl45叶片细胞中的绿色荧光信号极为明显(图 3g),表明在突变体spl45叶片中存在大量的PCD信号,能够触发植株叶片细胞发生细胞程序性死亡。
-
在抽穗期突变体spl45倒二叶叶片尖部刚出现斑点时,利用锡纸对未出现斑点的叶片部位进行遮光,WT也采取相应处理。7 d后去除锡纸拍照观察,发现WT遮光部分没有斑点产生而突变体spl45有少量斑点产生(图 4a)。复光一周后,WT未产生斑点,而突变体spl45迅速产生大量新的斑点且斑点遍布整个叶片(图 4b)。由此可见,突变体spl45叶片斑点产生受光照的影响。
-
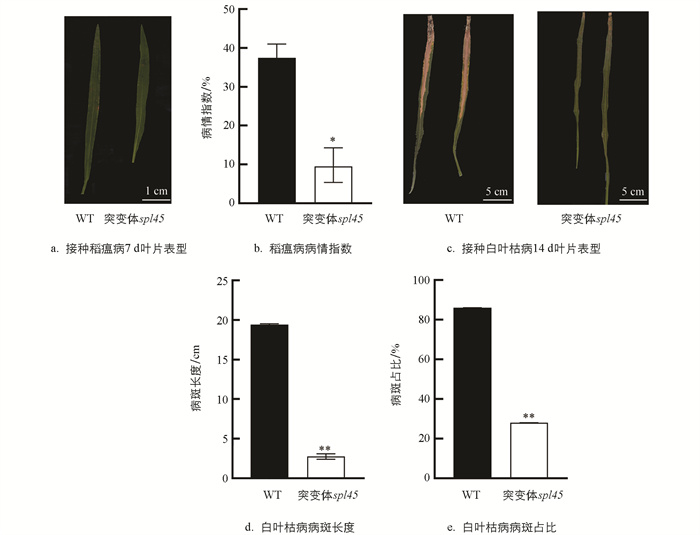
在大田中对WT和突变体spl45进行稻瘟病抗谱测定(表 4)。供试稻瘟病菌株划分为ZA、ZB、ZC、ZD共4个生理种群。苗期WT和突变体spl45接种稻瘟病菌7 d后待病情相对稳定时统计发病情况。结果显示,突变体spl45对稻瘟病的总群抗谱频率宽于WT,其对优势生理种群ZB、次优势生理种群ZC的抗病频率分别为100%、92.31%,对ZA、ZD两个生理种群的抗谱频率与WT无差异,表明突变体spl45的稻瘟病抗谱较WT有所拓宽。对WT和突变体spl45进行稻瘟病抗性鉴定,发现突变体spl45稻瘟病抗性显著提高(图 5a-b)。
孕穗期对WT和突变体spl45进行白叶枯病菌接种试验,接种14 d时调查其叶片病斑蔓延情况并记录病斑长度和病斑占叶片比,发现突变体spl45的白叶枯病菌病斑长度和病斑占叶片比显著低于WT(图 5c-e)。
综上,突变体spl45相较于WT,其稻瘟病抗谱拓宽,稻瘟病和白叶枯病抗性增强。
-
采用Jinhui 10号作为母本与突变体spl45作父本进行杂交获得F1,然后F1自交得到F2,共2 590株。本研究观察到在F2群体中有性状分离现象。突变体spl45基因的遗传分析如表 5所示,其中正常株为1 947株,突变株为643株,分离比为3.03∶1。通过卡方分析(χ2=0.68<χ0.052=3.84),卡方值远小于临界值,符合3∶1的理论分离比。由此可知,突变体spl45表型是受一对隐性基因控制。
-
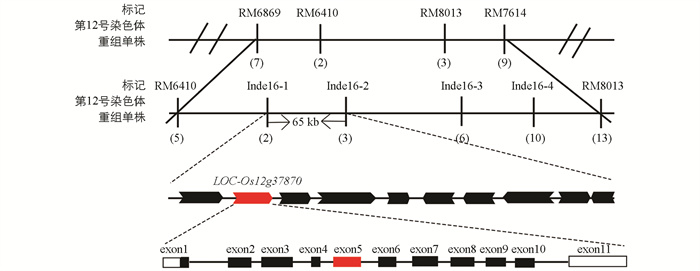
OsSPL45的基因定位如图 6所示。以突变体spl45与Jinhui 10杂交的F2群体作为材料进行基因定位,并使用均匀分布在水稻所有染色体上的SSR(Simple Sequence Repeats)标记引物进行差异性条带检验,发现在第12号染色体编号为RM6410和RM8013的分子标记之间存在显著的连锁效应。本研究在这两个分子标记之间设计4对InDel标记,并将目标基因定位于Indel6-1和Indel6-2之间的65 kb区间上。利用Gramene数据库(
http://www.gramene.org )和NCBI数据库(http://www.ncbi.nlm.nih.gov )来预测候选基因。经过序列检测发现,仅有编号为LOC_Os12g37870的基因编码区上第5个外显子经历了从碱基A到T的突变,导致其编码的氨基酸从天冬氨酸转变为缬氨酸,因此将LOC_Os12g37870作为OsSPL45的候选目的基因。 -
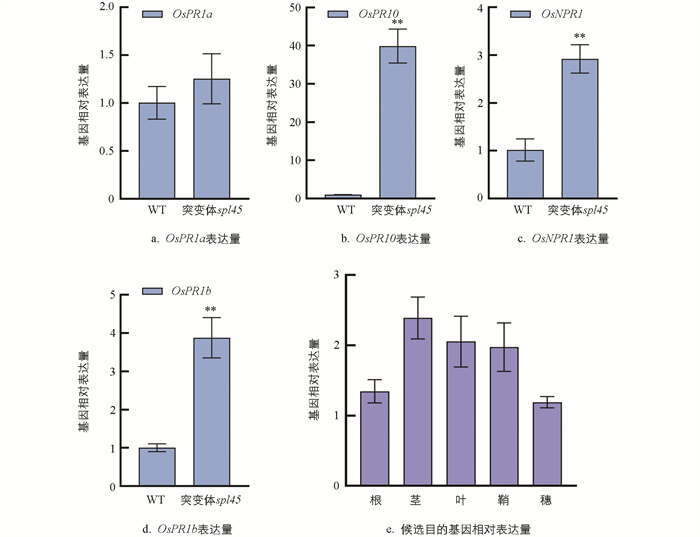
病程相关基因和目的候选基因相对表达量如图 7所示。通过qRT-PCR技术对分蘖期WT和突变体spl45叶片中病程相关基因进行检测,发现突变体spl45的OsPR1a、OsPR10、OsNPR1和OsPR1b表达量极显著高于WT,表明突变体spl45类病斑的出现可能触发了这些病程相关基因的过量表达,从而增强了对稻瘟病菌和白叶枯病菌的抵抗力(图 7a-d)。
对孕穗期WT根、茎、叶、鞘、穗候选目的基因的表达模式进行分析,结果显示OsSPL45在水稻各个部位均有表达,特别是在茎、叶、叶鞘中的表达量尤为显著(图 7e)。
2.1. WT和突变体spl45的表型鉴定
2.2. WT和突变体spl45的主要农艺性状分析
2.3. WT和突变体spl45的光合特性分析
2.4. 突变体spl45叶片的ROS积累与PCD分析
2.5. 突变体spl45类病斑表型受光照影响分析
2.6. 抗谱及抗性鉴定
2.7. 突变体spl45的遗传分析
2.8. OsSPL45的基因定位
2.9. 病程相关基因及候选目的基因的表达模式分析
-
根据出现的时间阶段,类病斑表型可以被分为3个不同类型,包括全生育期型、营养生长阶段起始型及生殖生长阶段起始型[21]。全生育期型指的是从突变体植物幼苗期一直到种子成熟的过程中,植物体持续出现有类病斑现象,而后两种类型则是在植株生长到某一特定阶段后出现类病斑表型。例如,lms1突变体在幼苗期便出现了褐色类病斑,这些类病斑会随着植株发育而逐渐扩散,最终在结实期斑点会遍布整个植株[22]。从突变体spl41的三叶期开始,植株叶片上开始逐渐出现红褐色斑点,然后叶片上的坏死斑数量和面积逐渐增加,导致整个叶片变黄并枯死[23]。像lmm7[24]和spl33[14]等突变体类病斑,在分蘖期才出现类病斑症状,并在后期生长阶段持续形成这类病斑。突变体spl45在幼苗期叶尖上就开始出现褐色斑点,随着植株生长和发育,这些坏死的褐色病斑数量逐渐增加,到了成熟期病斑已经遍布整个植株的叶片和叶鞘,表明spl45是一个典型的全生育期型水稻类病斑突变体。
大部分类病斑突变体的斑点会受到自然光的诱导且光合能力下降[12]。遮光实验及光合特性分析结果表明,突变体spl45上的类病斑受到光诱导,对自然光的利用能力减弱。类病斑突变体斑点部位及周围通常出现细胞程序性死亡,植株抗病能力有所转变的特点[25-27]。植物细胞PCD信号对植物有效免疫和防御病原体侵入具有至关重要的作用[28-29],通过诱导植物细胞死亡有助于限制病原体在入侵部位生长和增殖,从而提高自身抗病能力。已经报道的许多水稻类病斑突变体,如spl21、spl4、LIL1、sles、lmes6等发病部位常存在大量细胞程序性死亡,坏死病斑的部位及周围通常有活性氧过量积累,对某些常见水稻病害抗性显著增强[30-34]。本研究中突变体spl45表现出明显的细胞死亡现象,在叶片内有大量H2O2积累,清除酶稳态被破坏,对稻瘟病和白叶枯病具有广谱抗性,同时突变体spl45中部分防卫基因被激活,这些基因的表达量显著高于正常值。
近年来,水稻中与类病斑形成相关的基因相继被克隆出来,这些类病斑基因可编码不同类型的蛋白,包括转录调控因子、泛素连接酶、剪切因子、离子通道蛋白、蛋白激酶等[35]。例如,spl33编码蛋白可以结合不同的靶蛋白,从而介导不同的细胞凋亡过程[14, 36]。oscul3a突变体编码Cullin蛋白,Cullin3、RBX1和BTB蛋白组成RING型E3泛素连接酶,然后介导OsNPR1泛素化,从而负调控细胞死亡和免疫反应[37]。llb编码叶绿体甲基转移酶OsMTS1,可以直接或间接抑制油菜素内酯与茉莉酸的介导反应来影响水稻的防卫反应和生长应答[38]。本研究发现突变体spl45的变异表现受到一对隐性基因控制,经测序发现突变体spl45的LOC_Os12g37870基因编码区上第5外显子的碱基发生突变(A/T),编码氨基酸由天冬氨酸变为缬氨酸,导致翻译的蛋白质失去原有功能。OsSPL45与先前报道的OsSPL30[39]互为等位基因,但突变位点存在差异。据分析,OsSPL45异常编码一种ATP柠檬酸裂解酶(ATP-citrate lyase,ACL),该酶正常情况下在糖类的三羧酸循环(TCA)过程中催化柠檬酸和CoA生成草酰乙酸和乙酰CoA,其中乙酰CoA处于调节能量流动和物质循环的代谢中心,其分子合成对水稻生长和应激反应至关重要[40-42]。然而,OsSPL45具体如何影响TCA过程还需进一步研究。
-
突变体spl45是全生育期型受光诱导的水稻类病斑突变体,其三叶期开始出现褐色斑点类病斑,至分蘖期斑点分布于整个植株的叶片和叶鞘,并且该突变体在整个生长发育期都具有矮化和早衰表型。突变体spl45相较于WT光合色素含量降低,光合作用受阻,从而导致产量等农艺性状显著下降。突变体spl45的H2O2积累量在出现类病斑的部位相较于WT显著升高、PCD信号增强,POD、CAT、SOD活性较WT显著降低。突变体spl45叶片中抗性相关基因过量表达,对稻瘟病和白叶枯病的抗性相较于WT也显著增强。遗传分析表明,突变体spl45表型是由一对隐性核基因控制,且该基因定位于突变体的12号染色体上,通过测序把LOC-Os12g37870作为OsSPL45的候选目的基因。