



 下载:
下载:

-
开放科学(资源服务)标识码(OSID):

-
辣椒别名辣茄、海椒,是一种重要的蔬菜作物。辣椒病毒病在世界辣椒种植区均有发生,侵染辣椒的病毒种类在70种以上,我国报道约37种,如:黄瓜花叶病毒(cucumber mosaic virus,CMV)、烟草花叶病毒(tobacco mosaic virus,TMV)、番茄斑萎病毒(tomato spotted wilt virus,TSWV)、辣椒褪绿病毒(capsicum chlorosis virus,CaCV)、辣椒脉斑驳病毒(chilli veinal mottle virus,ChiVMV)等[1-5]。因气候和种植模式等影响,我国各地辣椒上的优势毒源种类有所不同,并不断发生着变化[6]。
辣椒是海南冬季重要的北运蔬菜,播种面积及产量均位居各类蔬菜之首,其中2022年播种面积为43 844 hm2,产量为108.14万t[7],主要种植在琼海、万宁、文昌、澄迈等地。病毒病是辣椒生产上危害最为严重的病害之一,特别是在长期连作种植区,严重影响了辣椒的产量和品质。本研究以一年生辣椒(Capsicum annuum)为研究对象,采用高通量测序结合RT-PCR检测,明确了海南辣椒病毒病的病原种类,分析了病毒复合侵染情况,并对CaCV的基因组分子特征进行分析,旨在为海南辣椒病毒病的科学防控提供理论依据。
全文HTML
-
2021年11月至2024年2月,从海南文昌、万宁、琼海、定安、临高、海口、澄迈和三亚共8个市县,采集疑似病毒病感染的一年生辣椒叶片样品184份。
植物总RNA提取试剂盒FastPure Universal Plant Total RNA Isolation Kit、反转录试剂盒HiScript Ⅱ 1st Strand cDNA Synthesis Kit、2×Rapid Taq Plus Master Mix和2×Phanta Flash Master Mix,南京诺唯赞生物科技股份有限公司;普通琼脂糖凝胶DNA回收试剂盒,天根生化科技(北京)有限公司;pMD 18-T Vector Cloning Kit、E.coli Competent Cell DH5α,宝日医生物技术(北京)有限公司。其他试剂均为国产或进口分析纯。
-
随机抽取16个辣椒叶片样品(每个市县2个样品),采用FastPure Universal Plant Total RNA Isolation Kit提取叶片总RNA,每8个样品的RNA等量混合为1个样本,共获得2个混合样本(LJ-POOL-1和LJ-POOL-2),送上海美吉生物医药科技有限公司完成深度测序。使用Hiseq 2000测序平台进行SE50测序,原始数据经过质控分析后,获得clean small RNA序列,使用Velvet 1.2.10软件对其进行拼接组装,将拼接结果在NCBI数据库中进行Blastn和Blastx比对,得到病毒序列进行统计并注释。
-
采用FastPure Universal Plant Total RNA Isolation Kit提取辣椒叶片总RNA,使用HiScript Ⅱ 1st Strand cDNA Synthesis Kit将提取的总RNA反转录成cDNA,使用病毒的特异性检测引物(表 1)对采集的184份样品进行RT-PCR检测,未标注参考文献的引物根据GenBank数据库中所登录的相应病毒基因组序列和高通量测序结果设计。PCR反应体系:12.5 μL 2×Rapid Taq Plus Master Mix、10 μmol/L的上游引物和下游引物各1 μL,1 μL cDNA,9.5 μL无菌ddH2O。PCR反应程序:95 ℃预变性2 min;95 ℃变性10 s,参照表 1退火温度退火30 s,72 ℃延伸10 s,循环30次;72 ℃延伸5 min。PCR产物经1%琼脂糖凝胶电泳检测,使用普通琼脂糖凝胶DNA回收试剂盒回收特异片段,回收产物直接测序。本研究中引物合成和测序均由北京六合华大基因科技有限公司完成。
-
根据田间样品检测结果,以辣椒样品WC24的总RNA为模板,基于高通量测序结果和GenBank已公布的CaCV基因组序列,利用在线软件Primer 3设计4对引物(表 1)扩增L、M和S RNA的主基因组序列,相应的5′和3′末端序列扩增使用Huang等[8]设计的引物。扩增获得的特异性片段产物经纯化回收后直接测序,或者将回收产物与pMD18-T Vector Cloning Kit连接,转化到E.coli Competent Cell DH5α中,利用PCR反应筛选阳性克隆。
-
测序结果采用NCBI中Blastn(
https://blast.ncbi.nlm.nih.gov/Blast.cgi )进行相似性查找。基于CaCV的L、M和S RNA基因组序列分别构建系统发育树,采用Clustal W法进行多序列比对,利用MEGA 11软件的邻接法(Neighbor-Joining,NJ)进行1 000次置信度自展分析,以西瓜银斑驳病毒(watermelon silver mottle virus,WSMoV)(GenBank登录号:U75379.1、AF133128.1和U78734.1)为外组。重组分析:使用RDP5软件提供的7种重组检测算法(RDP、GENECONV、BootScan、MaxChi、Chimaera、SiScan和3Seq)对辣椒褪绿病毒(CaCV)分离物的L、M和S RNA基因组片段序列进行潜在重组事件分析,每个重组事件需要至少被5种以上算法支持。
1.1. 材料来源
1.2. 方法
1.2.1. 高通量测序
1.2.2. RT-PCR检测
1.2.3. 辣椒褪绿病毒(CaCV)的基因组序列扩增
1.2.4. 序列分析
-
病毒病在海南辣椒主要种植区均有不同程度的发生,长期连作种植区发生率普遍高于新种植区。感染病毒病的辣椒在田间表现出多种症状,如叶片出现褪绿、丛簇、矮化、皱缩(扭曲)、花叶等症状,果实僵硬、斑驳、表面凹凸不平等(图 1)。
-
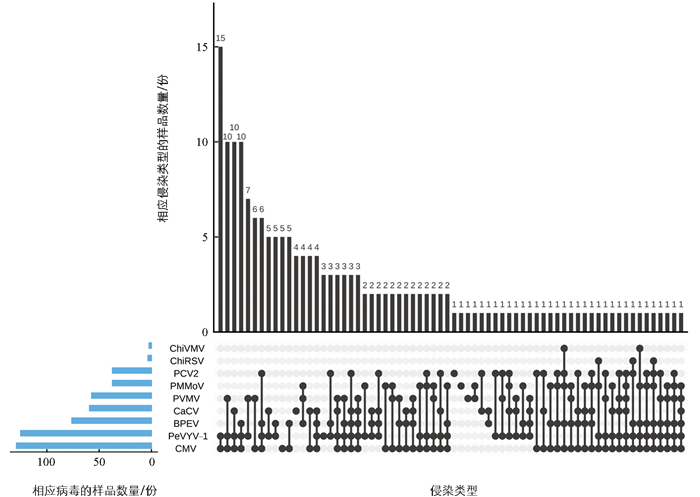
辣椒的2个样本(LJ-POOL-1和LJ-POOL-2)经高通量测序后共获得163 296 872条raw reads,质控后得到156 749 850条clean reads,其中74 619 976条reads比对到宿主参考基因组数据库,将未比对到辣椒参考基因组数据库的82 129 874条reads进行拼接组装,将拼接所得23 581条contigs与NCBI病毒库比对,筛选得到111条来源于植物病毒的contigs,其中BPEV 37条、CMV 20条、CaCV 19条、PCV2 9条、PMMoV 4条、PeVYV-1 20条、PVMV 2条。为了验证高通量测序结果的准确性,利用上述7种病毒的特异性检测引物(表 1)对高通量测序的样品进行RT-PCR检测,所有高通量测序分析出的病毒均得以确认。
-
前人研究表明,TMV、辣椒环斑病毒(chilli ringspot virus,ChiRSV)和ChiVMV在海南辣椒上发生普遍,但高通量测序并未发现这3种病毒。本研究在对采集的184份辣椒样品进行RT-PCR检测时,不仅检测到了高通量测序发现的7种病毒,还检测到了TMV、ChiRSV和ChiVMV。检测结果显示:6个样品未检测到病毒,病毒检出率为96.74%。CMV、PeVYV-1、BPEV、CaCV、PVMV、PCV2和PMMoV的检出率分别为70.11%、67.93%、41.30%、32.07%、30.98%、20.11%和20.11%,ChiRSV和ChiVMV检出率为1.63%和1.09%,未检测到TMV(表 2)。CMV、PeVYV-1、BPEV、CaCV和PVMV在所有市县的样品中均有检出;PCV2在除琼海和定安外的6个市县样品中均有检出;PMMoV在除临高和海口外的6个市县样品中均有检出;ChiRSV和ChiVMV仅在文昌和万宁的样品中有检出。PeVYV-1、BPEV、CaCV和PCV2是首次在海南辣椒上检测到的病毒。
在检测到病毒的178份样品中,仅15份样品为单一病毒侵染,其余样品为2~6种病毒复合侵染,复合侵染率为91.57%,有62种复合侵染类型(图 2),其中2种、3种、4种、5种和6种病毒复合侵染检出率分别为31.46%、29.21%、20.79%、7.87%和2.24%,说明海南一年生辣椒上病毒复合侵染现象严重,以2~4种病毒复合侵染为主。
-
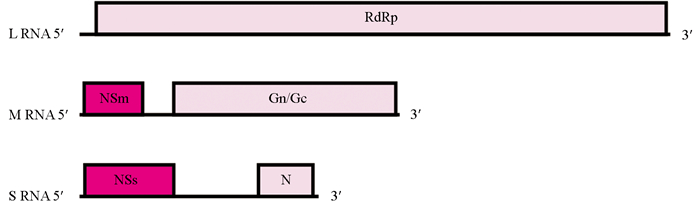
CaCV-WC24的L、M、S RNA的基因组长度分别为8 913 nt,4 840 nt和3 629 nt,GenBank登录号分别为PQ793445、PQ793446和PQ793447。CaCV-WC24的基因组结构见图 3,L RNA为负义,在互补链上含有1个开放阅读框(open reading frame,ORF),编码依赖于RNA聚合酶RdRp(248~8 881 nt,2 877 aa),分子量为331.20 kDa。M RNA为双义,含有2个ORFs,病毒链编码非结构蛋白NSm(58~984 nt,308 aa),分子量为34.32 kDa;互补链编码糖蛋白前体Gn/Gc(1 428~4 793 nt,1 121 aa),分子量为127.57 kDa。S RNA为双义,含有2个ORFs,病毒链编码非结构蛋白NSs(67~1 386 nt,439 aa),分子量为49.74 kDa;互补链编码核衣壳蛋白N(2 735~3 562 nt,275 aa),分子量为30.74 kDa。
Blastn比对显示,CaCV-WC24的L、M、S RNA基因组序列均与海南蜘蛛兰分离物(CaCV-Hainan)和藿香蓟分离物(CaCV-AC)的相似性最高(98.21%~99.27%),与其他CaCV分离物的相似性分别为83.98%~95.88%、92.21%~98.12%,87.80%~94.02%。CaCV-WC24的RdRp、NSm、Gn/Gc、NSs和N与已知CaCV分离物相对应基因的氨基酸序列相似性分别为93.67%~99.44%、91.56%~100.00%、91.44%~99.29%、87.93%~98.63%和94.91%~99.64%。
应用MEGA11软件,基于CaCV的L、M、S RNA的全基因组序列分别构建的3个系统发育树(图 4)。
由图 4可知,CaCV-WC24始终与另外2个来自海南的分离物(CaCV-Hainan和CaCV-AC)聚集在同一分支上。泰国番茄分离物CaCV-AIT在L和S RNA发育树中明显不同于其他CaCV分离物,单独聚在一个分支,而在M RNA发育树中,与其他8个分离物聚在一个分支。
基因组重组分析结果表明:CaCV-WC24的L、M、S RNA尚未发现重组事件,但CaCV-WC24的L RNA片段作为主要亲本参与CaCV-TN-CBE分离物(KT876916.1)的重组。
2.1. 辣椒病毒病的田间症状
2.2. 高通量测序结果及RT-PCR验证
2.3. 辣椒样品的病毒检测结果
2.4. 辣椒褪绿病毒(CaCV)的全基因组序列分析
-
病毒病是海南辣椒生产上危害最为严重的病害之一,科研人员在海南辣椒上检测到CMV、TMV、PMMoV、PVMV、ChiRSV、ChiVMV、芜菁花叶病毒(turnip mosaic virus,TuMV)、番茄黄化曲叶病毒(tomato yellow leaf curl virus,TYLCV)、蚕豆萎蔫病毒2号(broad bean wilt virus 2,BBWV2)、马铃薯Y病毒(potato virus Y,PVY)、黄瓜绿斑驳花叶病毒(cucumber green mottle mosaic virus,CGMMV)和番茄斑萎病毒(tomato spotted wilt virus,TSWV)共12种病毒[10-25]。本研究通过高通量测序和RT-PCR方法在海南一年生辣椒上发现了9种病毒:CMV、PeVYV-1、BPEV、CaCV、PVMV、PCV2、PMMoV、ChiRSV和ChiVMV,其中PeVYV-1、BPEV、CaCV和PCV2是首次在海南辣椒上发现,未检测到TMV。本研究检测到的病毒种类与前人的研究结果存在差异,可能是由于本研究采用的高通量测序技术可以一次性全面扫描样品中的所有病毒,而前人研究大多采用电子显微镜观察、血清学及RT-PCR技术,常局限于对已知病毒的检测;也可能是由于本研究检测对象为一年生辣椒(Capsicum annuum),而未包括在海南种植的中国辣椒(Capsicum chinense)和灌木状辣椒(Capsicum frutescens);或者是由于采样地点的不同,其栽培模式、自然环境的改变导致病毒种类发生改变。本研究中有6份疑似病毒病感染的样品未发现所检测的10种病毒,可能是样品中还存在其他病原性病毒,也可能是由其他病原菌或一些生理性原因所致。本研究样品中的病毒复合侵染率达到91.57%,病毒复合侵染高不仅增加了病毒病防控的难度,也极易导致抗病品种的抗性丧失,危害更为严重。
辣椒褪绿病毒(CaCV)最早报道于美国[26],随后澳大利亚、中国、印度、泰国、伊朗、希腊等相继有发生报道,可侵染辣椒(Capsicum sp.)、番茄(Solanum lycopersicum)、花生(Arachis hypogaea)、菠萝(Ananas comosus)、苦苣菜(Sonchus oleraceus)、万寿菊(Tagetes minuta)、马蹄莲(Zantedeschia spp.)、大萼球兰(Hoya calycina)、藿香蓟(Ageratum conyzoides)、飞机草(Chromolaena odorata)、朱顶红(Hippeastrum hybridum)、网球花(Scadoxus multiflorus)、蝴蝶兰(Phalaenopsis sp.)和一点红(Emilia sonchifolia)等多种植物[27-33]。海南在2017年就有CaCV侵染蜘蛛兰的报道,2023年,Sun等[34]在藿香蓟上也发现了CaCV的侵染,但一直未见关于CaCV侵染辣椒的报道。本研究通过高通量测序和RT-PCR扩增首次在海南辣椒上发现CaCV的侵染,并获得其全基因组结构。基于基因组序列的相似性比对和进化树分析发现,来自海南的CaCV分离物(CaCV-WC24、CaCV-Hainan、CaCV-AC)的基因组彼此间的相似性最高,而且始终聚集在同一进化分支上,说明海南CaCV的种群单一。
-
利用高通量测序及RT-PCR技术对来自海南8个市县的184份一年生辣椒样品进行检测鉴定,发现了9种病毒,其中CMV和PeVYV-1为优势病毒,检出率在67.00%以上,田间病毒的复合侵染现象普遍,新发现4种侵染海南一年生辣椒的病毒:PeVYV-1、BPEV、CaCV和PCV2。海南的辣椒分离物CaCV-WC24与蜘蛛兰分离物CaCV-Hainan和藿香蓟分离物CaCV-AC的基因组相似性最高,而且聚集在同一进化分支上。