


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
外来入侵植物通过迅速繁殖和排挤本地物种,导致区域生物多样性锐减,破坏了生态平衡,对生态系统构成了严重威胁[1]。外来入侵植物改变了入侵地的生态系统结构和功能,例如,假臭草(Praxelis clematidea)和马缨丹(Lantana camara)等物种在农田中的蔓延会影响土壤养分循环和微气候条件,导致农作物产量下降,增加农业管理成本[2]。此外,某些入侵植物通过根系释放化学物质,改变土壤结构和微环境,影响生态系统服务功能,如碳固定、氧气生产、水源涵养和土壤侵蚀控制等生态功能因入侵植物的蔓延而退化[3]。研究表明,入侵植物会通过竞争或占据本地物种生态位排挤本地物种,使本地物种的种类和数量减少,甚至导致物种濒危或灭绝,最终导致生态系统结构单一、功能退化,改变或破坏当地的自然景观。例如,加拿大一枝黄花(Solidago canadensis)的入侵导致上海已经有30多种土著物种局部消失[4];互花米草(Spartina alterniflora)入侵福建等地沿海滩涂,导致红树林湿地生态系统遭到破坏,原有的200多种生物减少到20多种[5]。目前退化生态系统正受到入侵植物的严重威胁,入侵植物的扩散对退化生态系统的稳定性、生物多样性和生态功能造成了极大的破坏[6]。入侵植物对本地物种生态位的侵占,已对退化生态系统中濒危植物造成了严重影响,阻碍了当地珍稀濒危物种保育工作的进行[7]。
银合欢(Leucaena leucocephala)又名白相思子、白合欢,为含羞草科多年生灌木或乔木,原产热带美洲,现广泛分布于世界热带、亚热带地区[8]。由于银合欢对引种地的乡土植物具有较强的抑制作用和化感效应,Lowe等[9]将其列为世界100种恶性外来入侵生物之一。一方面,银合欢适应性强,抗旱耐瘠、生长迅速,在一些生态退化区具有良好的水土保持和土壤改良等功能[10-11],近年来作为先锋树种,应用于我国金沙江干热河谷、西南石漠化区和采矿迹地等植被修复区域[12];另一方面,银合欢对生态环境的不利影响也已引起国内外有关方面的关注[13]。Ahmed等[14]研究了银合欢枯落物对黄豆树(Albizia procera)、豇豆(Vigna unguiculata)、鹰嘴豆(Cicer arietinum)和木豆(Cajanus cajan)等种子萌发和幼苗生长的抑制作用。陶文琴等[15]研究了银合欢叶水提液对芥菜(Brassica juncea)、菜心(Brassica parachinensis)和白菜(Brassica pekinensis) 3种植物种子萌发和根茎生长等的化感作用和特点。然而,银合欢对其他植物的影响不仅限于化感作用,其浓密的树冠结构可显著降低林下光照强度,从而与周边植物产生光合作用和生态位的竞争;同时,银合欢发达的根系也与周边植物产生激烈的水分、养分等资源竞争。近年来的野外观察发现,银合欢在我国西南石漠化区已出现一定程度蔓延,其冠层稀疏、凋落物量大的特性虽可在夏季减弱光照并改良土壤,但也通过遮荫和资源竞争抑制某些物种的生长。在桂西南石漠化区,以银合欢为优势的人工植物群落,普遍存在物种多样性低和群落层次单一等问题,群落生产力和生态服务功能较低,在物种组成和群落结构等方面需要加以人工促进或改造。尽管前期的研究表明银合欢在石漠化治理中的作用显著,但由于银合欢对一些植物或树种具有较强的抑制效应,因而在选择树种时需加以考虑和谨慎利用。
蒜头果(Malania oleifera)为我国特有单种属植物和国家二级重点保护植物[16],兼具水土保持、材用和油料等多重价值,属于珍稀濒危物种,在桂西南喀斯特地区广泛自然分布,表现出对石漠化生存环境的较强适应性。然而,在当前蒜头果直播造林过程中,普遍面临以银合欢为代表的外来入侵植物长期存在的生境保育、资源竞争、抢夺生态位等问题。这些入侵植物对蒜头果的保育与抑制作用之间的关系,目前尚未有详尽报道。此问题直接关系到蒜头果在石漠化地区直播造林项目的实际效果与成效评估。
本研究以西南典型石漠化区为研究对象,通过野外试验与盆栽试验,系统评价银合欢对蒜头果种子萌发和幼苗生长的影响,深入分析银合欢植株和不同枯落物(枯枝叶、干根等)对蒜头果的化感作用机制,以及蒜头果种质资源合理利用的管理策略,旨在为石漠化地区的植被恢复与可持续经营提供实践参考。本研究对于石漠化区造林过程中平衡外来物种的生态效益与潜在风险、保护本土珍稀物种、促进生物多样性和可持续造林实践具有重要意义。
全文HTML
-
试验分为野外直播试验(简称野外试验)和盆栽控制试验(简称盆栽试验)两部分,两者分别在广西平果市果化镇龙何屯喀斯特石漠化生态重建示范区(简称龙何示范区,23°23′N,107°23′E)和桂林植物园(25°01′N,110°17′E)进行。龙何示范区为比较典型的喀斯特峰丛洼地分布区,该区由于早期人为引种和后期的自然扩散,目前分布着一定数量的银合欢群落。龙何示范区毗邻右江河谷,属南亚热带季风气候,年均气温19.1~22 ℃,极端高温38.8 ℃,极端低温-1.3 ℃,大于等于10 ℃的年积温7 465.6 ℃;年降水量1 369.9 mm,5-8月降水量约占年降水量的70%,而9月至翌年4月降水量仅约占年降水量的30%,春旱和秋旱比较普遍和严重。桂林植物园位于广西桂林市郊雁山镇,属中亚热带季风气候,年均气温19.2 ℃,最冷月(1月)平均气温8.4 ℃,最热月(7月)平均气温28.4 ℃,极端高温40 ℃,极端低温-6 ℃,大于等于10 ℃的年积温5 955.3 ℃;年降水量1 865.7 mm,年均相对湿度78%。
-
试验地设在龙何示范区的中上坡,是自2005年起弃耕的荒坡地,坡向南,坡度22°~25°,土壤为棕色石灰土,以石窝土为主,盖度约30%,最大厚度不足35 cm。银合欢于2006年3月由人工定植并已形成一定规模的单优种群落,平均基径7.33 cm,平均株高3.05 m,平均冠幅2.08 m×2.19 m。冠下灌草植物高0.25~1.60 m,平均盖度约65%,主要有飞机草(Eupatorium odoratum)、蔓生莠竹(Microstegium vagans)、荩草(Arthraxon hispidus)、千里光(Senecio scandens)、肾蕨(Nephrolepis auriculata)、红背山麻杆(Alchornea trewioides)、假臭草(Eupatorium catarium)、类芦(Neyraudia reynaudiana)、九龙藤(Bauhinia aurea)和三叶鬼针草(Bidens pilosa)等。在播蒜头果种子前,以刈割方式全面清理除银合欢冠下的灌草植物,并在2012年5月至11月每月清理一次。
2013年3月下旬,将尚未萌发的蒜头果种子经层积沙藏(170 d)处理,筛出后穴播,播种穴深、穴宽均为20~25 cm,覆土深度4~5 cm,每穴点播种子1粒。试验包括2种处理:①播种穴位于银合欢冠下(YH),以单个石窝地为试验小区(面积3~4 m2);②播种穴位于冠外空地(由人工清除灌草植物而成,KD),以1~2个石窝地为试验小区(总面积3~4 m2)。每试验小区直播种子6~8粒,每处理8个重复。在试验观测期间,除定期割除灌草植物外,对蒜头果幼苗不进行培土和施肥。2013年6月下旬统计种子萌发数量并将幼苗挂牌标记,同年6月、9月和12月分别利用钢卷尺(精度0.1 cm)和数显游标卡尺(精度0.01 mm)测量各成活植株的株高和基径。
光照及土壤性状测定分别在2013年7月中旬和2013年11-12月进行。光照利用智慧型自动量程数字照度计(VICTOR1010A型,深圳市胜利高电子科技有限公司)于晴天的11:00-13:00,分别测定银合欢冠下及冠外空地距地面1 m处的光照强度,每个植株冠下及冠外空地(距离树冠外缘不小于1 m)各随机选择5~6个测点(取平均值,下同),以此求算树冠透光率。土壤性状测定包括物理性状和化学性状,前者采用100 cm3环刀法,主要测定土壤容重、总孔隙度和田间持水量等指标;后者主要测定pH值(电位法)、有机质含量(重铬酸钾滴定法-外加热法)、全氮含量(半微量开氏法)、全磷含量(钼锑抗比色法)和全钾含量(NaOH熔融火焰光度法)等指标。采集土样时,以单个石窝地为取样单元。物理性状测定:采用100 cm3环刀法于地表下0~1 cm处挖取环刀土样1个,用于测定土壤容重、总孔隙度和田间持水量等;化学性状测定:每个石窝地内随机选择3~4个位点,采集地表下1~15 cm深度的土壤,混合后取样约500 g,用于测定pH值、有机质含量、全氮含量、全磷含量、全钾含量等指标。
-
盆栽试验以圆形植树袋(直径80 cm、高60 cm)为装置,袋内装入从桂林植物园附近岩溶山地挖取的土壤(实测含水率25.03%,袋内土层高度约为43 cm),共设置5个处理:①每袋铺放风干的银合欢枯枝叶750 g(ZY);②每袋铺放银合欢干根1 500 g(GG);③每袋带土移植1株银合欢,平均株高2.1 m,平均基径3.43 cm(ZZ);④每袋移植1株银合欢并铺放银合欢枯枝叶750 g(模拟野外试验,ZK);⑤植树袋内既不移植银合欢植株,也不铺放银合欢枯枝叶或干根(空白对照,CK)。2015年1月17日至21日完成土壤装袋、银合欢移植,以及枯枝叶和干根的铺放(铺放深度为土层表面下方5 cm),每个处理5袋(试验重复)并南北排列成1行,行间距40 cm。2015年3月18日,以植树袋中心点为起点,在东、南、西、北4个方向至袋缘之间的中点处各穴播1粒蒜头果种子(层积沙藏约170 d),穴深4~5 cm,每袋4粒。从播种次日开始,每2 d观察和统计1次蒜头果种子发芽情况,种子发芽以胚芽出露0.5 cm及以上为准,直至连续5次(10 d)无新的种子发芽为止。6月22日,对蒜头果幼苗不足4株的植树袋,带土移植同时期播种、大小相近的袋装苗,使每袋幼苗总数达到4株。从2015年3月至12月,除正常浇水和除草外,对蒜头果幼苗不进行松土和施肥,每月下旬(11月除外)分别利用钢卷尺和数显游标卡尺测量各成活植株的株高和基径。2015年12月17日,利用便携式叶绿素测定仪(型号SPAD-502,精度0.1)测定叶片SPAD值,即在每个成活植株上、中、下部位各选取1个形态正常、无病虫害的成熟叶片,每个叶片随机测定5~6个位点并取平均值,然后将叶片剪下带回实验室,用便携式叶面积仪(型号Li-3000A,精度0.01 cm2)逐一测定单个叶片面积,并在70 ℃烘箱内烘干48 h后用电子分析天平(METTLER-AE240,精度0.000 1 g)称取单个叶片重量,以此求算比叶重(mg/cm2)。
-
为分析和比较银合欢的植株、干根和枯枝叶等对蒜头果幼苗生长的影响,采用相对相邻作用系数(RII)[17],RII的计算公式为:
式中:XN为各处理(YH、GG、ZY、ZZ和ZK)中蒜头果幼苗的平均幼苗保存率、株高或基径值;X0为相应对照组的平均值;RII取值范围为[-1,+1],RII>0表示相对相邻作用是正效应,即银合欢植株、枯枝叶和干根等对蒜头果幼苗生长具有促进作用;RII<0表示相对相邻作用是负效应,即银合欢植株、枯枝叶和干根等对蒜头果幼苗生长存在抑制作用。RII绝对值越大,表明银合欢植株、枯枝叶和干根等的促进作用或抑制作用越明显。
数据分析采用SPSS 19.0软件,绘图采用Origin 2021完成。使用单因素方差分析,分别比较野外试验和盆栽试验之间的差异,检验使用LSD显著性检验,p=0.05。
1.1. 研究区概况
1.2. 试验设计与观察
1.2.1. 野外试验
1.2.2. 盆栽试验
1.3. 数据分析
-
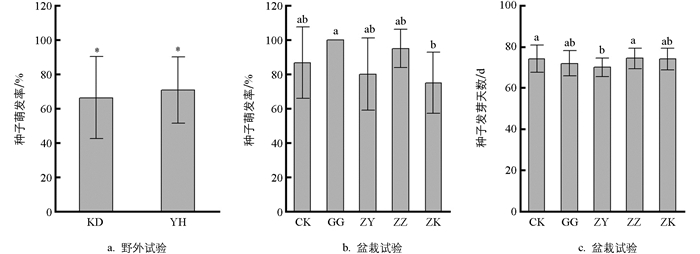
据测定(表 1),银合欢冠下(YH)平均透光率为58.91%,与冠外空地(KD)差异有统计学意义(p<0.01),表明在光照强烈的夏季,银合欢树冠能够显著降低冠下光照强度。在土壤性状方面,银合欢冠下与冠外空地在土壤容重、总孔隙度、pH值、有机质含量、全氮含量、全钾含量和全磷含量指标上差异均无统计学意义,仅田间持水量在冠下显著高于冠外空地。表明银合欢对冠下土壤的物理性状能起到一定的改善作用,但对化学性状的改善作用较小。
-
野外试验(图 1a)种子萌发率平均值分别为66.3%(KD)和70.7%(YH),不同处理之间差异有统计学意义。盆栽试验(图 1b)在播种后第64 d(5月20日)种子开始萌发,至第84 d(6月9日)结束,持续时间约20 d,种子萌发率平均值从大到小依次为GG、ZZ、CK、ZK、ZY,但除了ZK的萌发率显著小于GG(p<0.05)外,其他不同处理之间差异无统计学意义。种子平均萌发天数为70~74.3 d(图 1c),其中ZY与CK,ZY与ZZ之间差异有统计学意义(p<0.05),其他处理之间差异无统计学意义(p>0.05)。
-
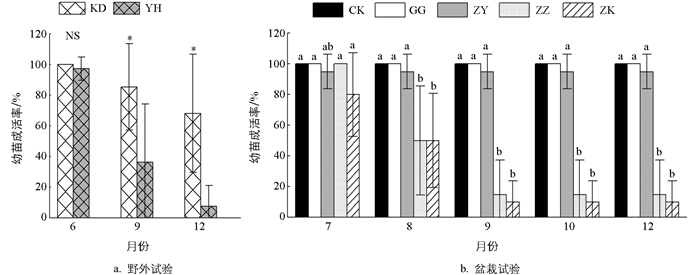
2013年6月、9月、12月,野外试验(图 2a)的幼苗成活率持续下降。其中2013年9月和12月不同处理之间差异有统计学意义(p<0.05)。与冠外空地(KD)相比,银合欢冠下(YH)的幼苗死亡率更高,其在2013年9月和12月的成活率分别仅为36.2%和7.6%,而冠外空地则达到85.3%和65.1%。在盆栽试验(图 2b)中,2015年7月至12月,GG、ZY和CK的幼苗成活率基本上保持不变,平均值均在95%以上且两两之间差异无统计学意义。而ZZ和ZK则持续下降,2015年12月时分别仅为15%和10%,两者从8月至12月与GG、ZY和CK的差异均有统计学意义(p<0.05)。观察发现,盆栽试验幼苗死亡最早发生在6月23日(ZK),而死亡高峰期在9月上、中旬。试验结果表明,在有银合欢植株的试验处理中,蒜头果幼苗死亡率很高,而其他试验处理的死亡率较低,说明银合欢植株对蒜头果幼苗具有比较强烈的抑制作用。
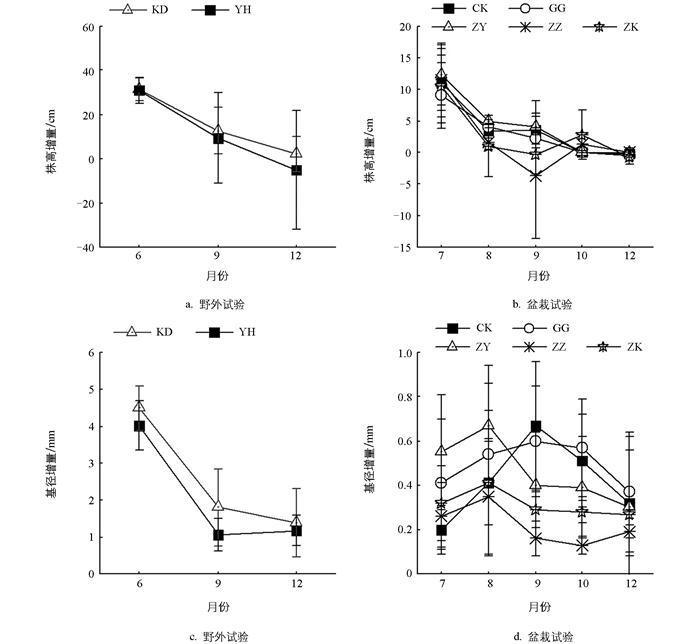
2013年6月至12月,野外试验的幼苗株高增量和基径增量逐渐下降,但均表现出KD大于YH的特点,其中10-12月YH的株高增量为负值,主要原因是冠下存活幼苗均发生不同程度的枯梢。在盆栽试验中,2015年6月至12月,CK、GG和ZY 3个处理的株高增量逐渐下降,而基径增量最大值出现在8月(ZY)和9月(CK和GG)。ZZ和ZY的株高增量从7月到9月逐渐下降,10月有所回升并在11月至12月显著下降,基径增量最大值出现在8月,而9月至12月逐渐下降,主要是两个处理在8月至9月幼苗死亡较多,且存活植株均出现枯梢,而10月份存活植株均从主干萌生直立新枝,使株高增量表现为正值(图 3)。
-
在野外试验中,银合欢冠下(YH)蒜头果幼苗的平均株高、平均基径均小于冠外空地(KD)且差异有统计学意义(p<0.05)(表 2),其相对相邻作用系数(RII)分别为-0.20和-0.33,即银合欢对蒜头果幼苗生长表现为负效应。在盆栽试验中,GG和ZY处理蒜头果幼苗的平均株高和平均基径均大于CK,其中ZY的平均株高与CK差异有统计学意义(p<0.05);ZZ和ZK的平均株高和平均基径均小于CK,两者平均株高以及ZZ的平均基径与CK的差异有统计学意义(p<0.05)。RII值计算结果显示,银合欢干根和枯枝叶对蒜头果幼苗的相对相邻作用为正效应(RII>0),而银合欢植株则为负效应(RII<0)。此外,GG和ZY的叶片SPAD值和单叶面积均大于CK,其中ZY的叶片SPAD值、单叶面积与CK的差异均有统计学意义(p<0.05),GG和ZY的比叶重小于CK且相互之间差异有统计学意义(p<0.05)。
2.1. 银合欢冠下和冠外空地的光照及土壤性状特点
2.2. 种子萌发
2.3. 幼苗成活与生长
2.4. 幼苗性状
-
植物相邻个体之间的相互作用是影响植物群落结构的重要因素[18-19]。对不同层次的植物而言,上层植物对下层植物的作用主要表现为保育促进、竞争抑制和中性共存3种形式[20-21]。从已有的研究结果来看,入侵植物对其他植物(目标植物)有无影响主要与入侵植物本身特性[22]、周围环境条件[23-24]、目标植物特性及其生长阶段[25-26]等相关。本研究发现银合欢对蒜头果种子萌发率和幼苗不同阶段的存活率产生一定影响,这一现象体现了银合欢对于蒜头果生命早期作用的复杂性和动态性。具体来说,银合欢的活体植株、枯枝叶和干根对蒜头果种子萌发的直接影响相对有限,这可能是因为蒜头果在种子萌发阶段根系分布范围具有局限性,对化感物质的敏感度较低,也可能是银合欢早期移植阶段释放的化感物质种类和浓度不足以对蒜头果种子萌发产生显著抑制作用[27]。然而,当蒜头果进入幼苗生长期后,银合欢植株通过根系分泌物或叶面淋溶物质对蒜头果幼苗展现出明显的抑制作用,这表明此阶段银合欢可能释放了对幼苗生长特别敏感的化感物质;相比之下,银合欢的枯枝叶和干根通过分解为土壤提供额外营养,在一定程度上促进了蒜头果幼苗的生长。这反映了化感作用的双重性质——既可以是竞争和抑制,也可以是资源供给和促进。Zhang等[28]的研究进一步支持了上述观点,他们发现桉树(Eucalyptus sp.)的根桩和枯落物对锥(Castanopsis chinensis)、山杜英(Elaeocarpus sylvestris)和枫香(Liquidambar formosana)等树种的影响各异,说明化感作用的效果高度依赖于作用对象的特性和所处的生命阶段,即使同一物种产生的化感物质,也可能对种子萌发和幼苗生长产生正、负不同的效应,这强调了在生态管理和直播造林实践中需要细致考虑化感作用的多维度影响。银合欢在蒜头果萌发早期阶段微环境中的化感作用并非单一方向,而是表现为化感物质释放模式、作用机制和浓度依赖性的复杂组合,其对生态系统长期健康与生物多样性的影响也随目标植物种类和生命阶段的不同而不同。
相关实践证明,外来入侵植物是退化生态系统和其他极端生境中常见的先锋植物,但入侵植物存在释放化感物质和过度竞争资源的特性,不利于群落的演替[29-31]。植物间的化感作用是生态系统内物种间相互作用的重要方面,对于理解植物群落结构的形成、物种的共存机制具有重要意义[18]。化感物质的释放途径多样,包括但不限于地上部分的挥发、雨雾淋溶作用下从叶片表面释放的化学物质,以及根系直接向土壤中分泌的物质[32]。植物凋落物的分解过程也是化感物质释放的关键途径之一,这些物质可被土壤微生物转化或直接作用于周围植物[28]。本研究发现银合欢植株对蒜头果幼苗成活与生长具有显著抑制效应。银合欢的这种抑制作用很可能与其根系分泌物密切相关。王仙等[33]的研究发现油松根系浸提液对不同植物发芽和生长的影响具有复杂性,油松根系分泌物在低浓度时对油松、胡枝子和绣线菊等植物具有一定的促进作用,但随着浓度增加,转而表现为抑制作用。类似地,银合欢的根系分泌物可能含有特定的化感物质,这些物质在土壤中积累到一定浓度时,能够显著抑制蒜头果幼苗的生长,从而影响其成活率和生长速度[8]。此外,除化感作用外,银合欢还表现出很强的资源竞争能力。银合欢的树冠结构对蒜头果幼苗的间接影响也不容忽视,其冠层结构密集,透光率较低,显著影响林下光照条件,进而抑制冠下喜光植物的光合作用和幼苗生长[11]。尽管如此,这种树冠遮阴效果在某些情况下有利于土壤水分保持,减少水分蒸发。同时,其枯落物的积累和分解有助于改善土壤结构和提高土壤肥力,为其他植物生长创造有利条件[26]。
综上所述,银合欢对蒜头果幼苗的抑制作用是多因素共同作用的结果。尽管本研究初步通过盆栽试验在光照一致条件下发现银合欢单种处理仍显著抑制蒜头果生长,排除了单一树冠遮光因素的干扰,但尚不能完全排除其他环境因子的影响。此外,枯枝叶与干根处理未表现出明显抑制效应,反向说明抑制效应可能源于活体根系分泌的潜在化感物质。因此,在生态恢复实践中,应权衡银合欢带来的正面效应(如土壤改良和水土保持)与其对特定本地物种产生的抑制作用。建议通过控制银合欢种植密度、引入多样化物种组合,或在银合欢生长特定阶段进行适度移除,以减轻其对本土植物的潜在负面影响[29]。通过这种综合管理,可以更有效地利用银合欢的生态功能,同时最小化其对脆弱生态平衡的干扰,推动生态恢复。
-
本研究揭示了银合欢对蒜头果的复合影响机制。研究结果表明:银合欢植株显著抑制蒜头果幼苗成活与生长,野外冠下幼苗成活率显著低于冠外,其抑制作用主要源于冠层遮光导致的光合作用受限和根系对水养资源的竞争,潜在化感物质也参与调控。因此,在石漠化区植被恢复中,建议蒜头果造林严格规避银合欢冠层覆盖区,优先选择林窗、林缘等光照充足、竞争较小的生境,并可探索凋落物土壤改良潜力与活体植株抑制风险的平衡管理。本研究可为喀斯特脆弱生态系统修复树种优化配置及区域生态安全维护提供科学依据。