


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
人类的面孔可传达个体身份信息,同时承载着情绪、注意力以及意图等各类社会信号,面孔加工被视作一种人类社会互动以及心理发展的基础性认知能力[1-2]。人类在进行面孔识别时会呈现出极为敏感且快速的加工能力,大量心理学研究显示,婴儿在出生之后的数小时之内便可快速对面孔样式的刺激产生偏好[3],这种早期偏好为后续的社会知觉、归因以及情绪理解奠定了认知基础[4]。对于成年人,面孔识别具备高度自动化的加工机制,体现了更快的速度及准确性,涉及对熟悉性、情绪状态以及个体差异的精细辨别[5-6]。面孔识别中的重复抑制(Repetition Suppression,RS)被解释为神经加工效率的提高,大脑可借助表征“精炼”[7-8]或者预测机制[9-10]减少对重复信息的资源消耗。通过功能核磁共振成像(Functional Magnetic Resonance Imaging,fMRI)等神经影像技术可观察到,当相同面孔被重复呈现的时候,大脑对于该刺激的反应往往会减弱[11]。由于面孔识别属于复杂的感知-认知进程[12],而RS又是快速加工该进程的一种具体体现,因此理解RS将为揭示面孔识别的神经加工机理提供一种重要研究手段。
虽然在认知心理学中RS是被广泛观察到的一种现象,但关于其神经机制的研究仍在不断发展和变化,尤其在面孔加工通路里,RS是依赖脑区内的活动还是区域间连接的调制仍值得探讨。早期的RS研究大多聚焦于单一区域的激活变化,但随着大脑网络模型的兴起,越来越多的研究开始转向探讨脑区间连接的动态变化。在面孔加工的分层神经系统中,右枕叶面孔区(rOFA)和右梭状回面孔区(rFFA)被视为2个关键感兴趣区(ROI),分别承担面孔的结构性分析与身份识别加工[13]。原先的观点强调RS更多是OFA或FFA等单一区域反应变化的体现,但随着动态因果建模(Dynamic Causal Modeling,DCM)等[14]的发展,越来越多的研究开始将重点放在区域间有效连接的调制机制上。重复刺激引发的神经变化不仅表现为局部活动的变化,还可能表现为信息在神经通路上的变化。
FI模型是较早对面孔加工网络的有效连接进行研究的一种模型,着重突出了rOFA、rFFA与顶叶、前额叶等高级区域间的动态交互作用,认为加工网络并非线性层级结构,而是具有分布式、多通道的特性[15]。这一观点扩展了传统“自下而上”加工路径的认知,为网络层级方面的建模奠定了基础。随后的研究在方法学层面对FI模型进行了反思与再验证,指出在不同实验设计及不同模型结构的情况下,有效连接的估计结果可能出现变化[16],应当依据具体范式重新审视面孔加工网络的“有效连接性”,而不是把某一连接方向当作固定加工通路。这一主张和RS机制的多样性高度契合,为理解不同重复面孔条件下的连接调制机制提供了关键的方法学提示。
在具体研究不同面孔重复条件对RS的影响中,当实验设计为隐式任务(即被试并未有意识地对面孔展开辨别)时发现,立即重复主要提高rFFA→rOFA的反馈连接,而延迟重复则调制rOFA→rFFA的前馈路径[10],这提示了时间结构或许是调制方向性的关键因素。但需注意,该发现采用的是“重复类型组合对比”策略,难以揭示单一重复条件的独立连接效应。此外,还有研究发现面孔的熟悉度显著影响隐式任务下的RS效应,只有熟悉面孔的重复才引发显著激活下降,陌生面孔则无此效应[17-18]。尽管这些研究仍聚焦于局部活动变化,但从熟悉度的角度对连接层面提出了潜在启发。
由上所述,RS可能凭借提高或者抑制特定方向的连接来调制信息加工流,但其方向性却存在差异,且依赖于熟悉度。这表明RS可能并非由单一机制所驱动,而是受到多重因素的调控,包括“隐式vs.显式”[18-19]、“立即面孔vs.延迟面孔”(时间结构)[10]、“熟悉面孔vs.陌生面孔”(语义属性)[20]以及“面孔vs.打乱面孔”(完整感知)[10]等。鉴于这些差异,当前仍然不清楚以下2个问题:①面孔的重复时间结构是否会与语义属性、完整感知共同发挥作用,调制特定方向的有效连接?②在隐式任务范式下,不同重复类型是否在连接层面呈现出各自独立且可区分的调制效应?
此外,现有的实验设计大多凭借组合对比(如不同语义属性与不同时间结构的组合)间接推断连接调制模式,很难识别各重复条件自身的调制特征。因此,本研究采用fMRI数据,尝试引入更为精细的建模策略,从单一重复条件入手,剖析重复时间结构、语义属性以及完整感知的交互作用机制,以期揭示重复抑制在连接层面的具体实现方式。
全文HTML
-
本研究采用开源神经数据库OpenNeuro中的数据集“A multi-subject,multi-modal human neuroimaging dataset”[10, 21-22],具体网址为
https://openneuro.org/datasets/ds000117/versions/1.1.0 。该数据集为面孔识别范式下的多模态脑影像数据(M/EEG和fMRI数据),本研究采用了其中的标准化fMRI数据,其由Siemens 3T TIM Trio MRI设备所采集。先前版本的数据包括19名健康成人(年龄范围23~39岁,有8名女性),为确保数据的完整性,经多次数据筛选,最新版本的数据集中仅保留了16名受试者的数据。结构像为T1加权高分辨率图像,功能像则通过梯度回波EPI序列采集,扫描参数为:TR=2 000 ms,TE=30 ms,空间分辨率3×3×3.75 mm,横断面切片32个。每位被试完成9个功能性fMRI runs,数据集中还提供了详细的刺激时序、反应按键信息以及头动参数等辅助数据,便于后续分析中的建模与共变量控制。数据采集过程中,所采用的面孔识别范式是让初次和重复刺激以伪随机方式交替呈现,规避了预期抑制的影响[9]。刺激的面孔具有不同的类型,包括熟悉面孔、陌生面孔和打乱面孔。面孔刺激包括2组共计300张灰度照片,一半的面孔为男性,另一半为女性。名人面孔(熟悉面孔)的选择旨在确保大多数成年人都能识别,非名人面孔(陌生面孔)在性别和年龄上与名人面孔大致匹配(通过视觉判断实现)。此外,每张打乱面孔图像都是从对应编号的名人或非名人面孔生成的,打乱方式是对原始面孔进行二维傅里叶变换,打乱其相位信息后再进行反变换恢复图像。为了让打乱图像在整体形状和大小上近似于原始面孔,这些图像被进一步裁剪为由1张名人和1张非名人面孔组合生成的掩模形状。虽然这种掩模处理导致打乱图像的功率密度谱不再与原始图像完全一致,但研究者认为匹配2类刺激在视觉角度上的一致性更加重要,例如最小化眼动差异。所有照片都经过统一处理和裁剪,仅显示面部区域,照片同时展示了多种发型(但长发已被裁剪)、表情(以快乐或中性为主)和角度(均为正面至3/4视角之间)。这2组照片中,一组照片用于M/EEG采集,另一组用于fMRI采集。
实验的设计详见文献[23],刺激材料通过投影仪呈现在距离参与者大约1.3 m远的屏幕上,水平方向和垂直方向的视角分别约为3.66°和5.38°。照片呈现在黑色背景上,中央有1个白色注视十字。每次测试的开始以1个注视十字的出现为标志,持续时间在400~600 ms之间随机变化。之后,关键刺激(面孔或打乱面孔)呈现,持续时间在800~1 000 ms之间随机变化。刺激呈现前的随机延迟是为了减少对正在进行的神经振荡(尤其是5 Hz以上频率)的混叠效应,并避免刺激前的相位重置。刺激结束时的随机延迟用于平均化视觉偏移带来的影响(同样针对5 Hz以上的频率)。2个刺激之间的间隔呈现1个中心白色圆圈,持续1 700 ms。要求参与者在整个实验过程中注视屏幕中心,中心圆圈到中心十字的变化旨在帮助参与者准备接收接下来的刺激,他们还被指示在注视十字或刺激呈现期间尽量不要眨眼(但在出现白圈时可自由眨眼)。
每张图像会呈现2次,第2次呈现的时间可能紧接第1次之后(立即重复),也可能是在5~15个其他刺激之后(延迟重复),这2种重复各占50%。设置重复间隔的目的,是因为已有研究表明该因素会影响重复相关效应[24]。综上,所有的测试包含3种面孔类型:熟悉面孔、陌生面孔、打乱面孔,以及3种刺激类型:初次刺激、立即重复和延迟重复。
为确保参与者注意每一个刺激,他们被要求用左右食指中的一个按下两个按键之一(按键分配在被试之间进行平衡),按键选择依据是参与者主观判断该图像“是否比平均对称程度更高或更低”。对称程度的平均标准通过练习环节来建立,练习中呈现了23张与正式实验无关的照片,参与者需重复练习,直到熟悉该任务。选择该任务的理由是其适用于面孔和非面孔图像[23],但由于该任务的对称性判断具有一定主观性,因此行为数据中没有特别感兴趣的内容(除了平均反应时,其均值约为955 ms,标准差约为283 ms)。需要指出的是,被试未被告知是否需要识别面孔身份或者判断刺激是否重复,而是执行一个与面孔加工没有直接关联的任务——判断面孔是否对称。在这种任务设计中,由于被试并不需要判断面孔是否重复,只是被动接收面孔刺激,因此该范式可被归类为隐式任务范式,避免了认知策略对重复效应的显性干预,使实验更贴近日常面孔识别过程中的无意识加工,可剖析重复抑制效应下的自动性神经机制。
-
本研究采用SPM12(
www.fil.ion.ucl.ac.uk/spm )软件工具箱进行fMRI数据的预处理,所有被试的预处理均依据统一参数完成,以保证后续分析具有一致性与可比性,具体步骤如下:①头动校正——移除每位被试每阶段起始的2个时间点图像,把其余图像配准至第1个时间点,对头动给予修正并生成平均图像,接着完成重采样来提升空间一致性。②时间校正——针对有33层交错采样切片的功能图像,将TR设置为2 s,把参考切片设定为第2层,运用统计参数图(Statistical Parametric Mapping,SPM)的spm_slice_timing函数来进行切片时序校正。③结构像与功能像配准——借助spm_coreg与spm_get_space函数,把T1结构图像借助刚体配准到平均功能图像上,以此达成空间一致性。④组织分割与标准化参数估计——采用spm_preproc与spm_preproc_write对结构图实施灰质、白质以及脑脊液的分割,并生成标准化参数。⑤空间标准化——利用上述参数把结构图和功能图分别归一化到MNI305空间,分辨率分别设置为1×1×1 mm和3×3×3 mm。⑥空间平滑——对标准化后的功能图像应用FWHM为8 mm的三维高斯核进行平滑,提升信噪比并契合随机场理论的统计假设。虽已有研究显示右上颞沟(rSTS)等区域也参与面孔信息加工,但因本研究聚焦面孔重复范式下的知觉加工效应,选取rOFA和rFFA 2个区域作为建模核心更合适。一方面,rOFA和rFFA在面孔视觉通路里分别承担初级特征分析与个体身份识别的关键作用,是重复效应最直接的传递通道[13, 25];另一方面,由面孔识别任务的DCM研究说明,rSTS未显示出强的重复调制作用,其包含在模型中可能引入冗余连接,降低模型稳定性[10]。基于此考虑,最终在模型设计中排除rSTS等与重复调制关联不大的高阶区域,仅聚焦rOFA与rFFA 2个核心区域,兼顾模型结构简洁性、参数估计稳定性以及调制效应的明确性。
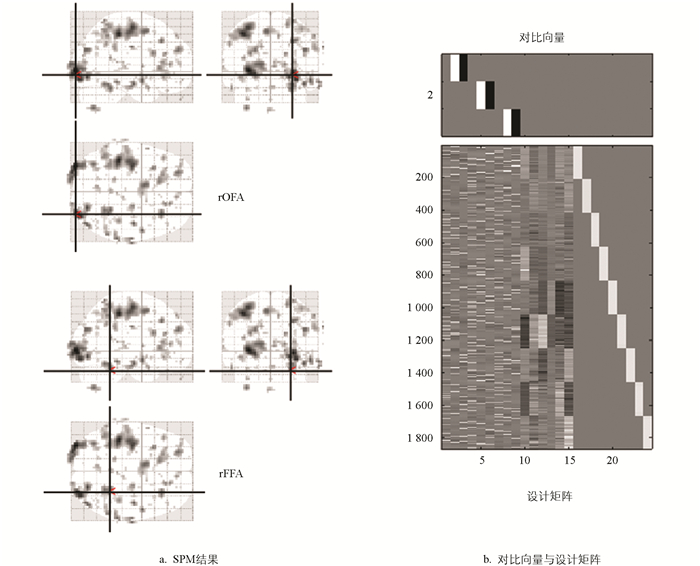
在ROI的定位上,与传统基于解剖模板(如AAL)和基于固定坐标点的策略不一样,本研究采用了模板引导下的功能定位方式来选取ROI。该定位方式把结构一致性与任务相关激活的个体差异性相结合,可提升ROI提取的功能特异性以及跨被试的稳定性。先参考AAL模板里rOFA与rFFA的结构位置,以此作为初步的空间限定范围。在一级广义线性模型(GLM)[26]分析中,定义9个回归因子,分别为:①初始熟悉面孔;②立即重复熟悉面孔;③延迟重复熟悉面孔;④初始陌生面孔;⑤立即重复陌生面孔;⑥延迟重复陌生面孔;⑦初始打乱面孔;⑧立即重复打乱面孔;⑨延迟重复打乱面孔。据此,构建一个包含3组条件对比的F对比矩阵,即:
矩阵中的3行分别对应熟悉面孔、陌生面孔与打乱面孔在立即重复与延迟重复条件下的激活差异。F对比可呈现面孔重复类型对皮层响应的整体调制效应,为功能定位提供关键依据。根据F对比生成的激活图,结合AAL模板的结构掩模,在可识别激活的区域之内对候选ROI坐标进行人工确认以及微调。
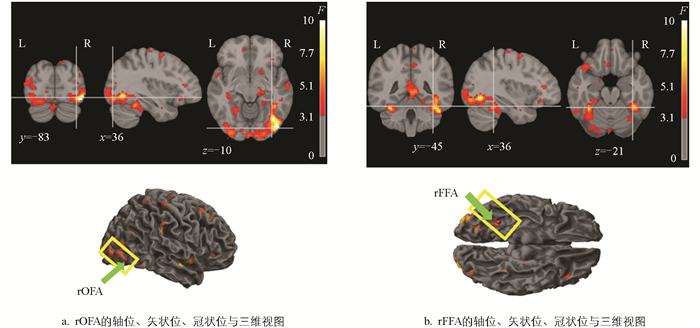
图 1给出了由上述F对比生成的一个被试的具体激活图,其中阈值设定为p<0.05,未校正,最小簇大于10。
激活图里呈现出的分布模式,与AAL模板中rOFA和rFFA的解剖位置极为相符,这证实了本研究所构建的F对比在确定面孔重复加工路径中任务相关脑区的有效性。另外,为了补充激活的统计特征,列出了其中一个被试在该对比结果中与rOFA和rFFA区域相关的激活峰值坐标、统计值以及簇大小,如表 1所示。
虽然rOFA和rFFA在个体上的功能具有高度一致性,但空间位置可能会出现变异[15],因此表 1中的坐标仅是计算结果,并不是最终结果。为防止“标准空间”坐标在个体脑图中的偏移造成信号提取偏差,采用数据驱动的点位验证策略提取信号,其步骤如下:①依据图 1中SPM软件的统计模型结果,调用spm_results_ui和spm_list函数,提取各被试的较大激活点坐标,并保存到统一的数据结构中。②依据rOFA和rFFA的MNI坐标范围,挑选位于这些空间范围内各被试的较大激活点坐标作为候选ROI激活点集合,具体的坐标范围为:rOFA的x∈[30, 40],y∈[-95, -70],z∈[-20, 0];rFFA的x∈[30, 40],y∈[-65, -35],z∈[-25, -5]。③采用“最大激活点覆盖”策略,在每个ROI预定义的空间范围内,以2 mm为步长生成候选中心点;对于每一个候选点,计算其在半径10 mm的球形区域内所包含的激活点数量,将覆盖激活点数最多的候选点作为该ROI的最优中心坐标。④以最优中心坐标为球心,构建直径为10 mm的球形区域,提取球形区域内所有体素的第一个主成分作为对应ROI的信号。
为验证ROI半径选择的稳健性,本研究进行了敏感性分析。在保持ROI中心不变的前提下,将球半径扩大至12 mm,重新提取BOLD时间序列并拟合DCM。为确保ROI提取的BOLD信号具有空间有效性,引入了明确的被试纳入标准,即如果在某一被试中,依据最优坐标构建的任一ROI(rOFA或rFFA)掩模内不包含有效体素(即掩模体素数为0),则认为该被试在该ROI上无法获得可靠的信号。
为评判由ROI坐标所提取的BOLD信号序列是否可用于DCM分析,可以通过DCM模型拟合出被试的一个ROI的BOLD信号,并计算该BOLD信号的方差解释比例,定义如下:
式中:Y为真实BOLD信号;
$ {\mathop {\boldsymbol{Y}}\limits^ \wedge}$ 为DCM拟合的BOLD信号;μ为真实BOLD信号的均值。VE指标衡量了模型解释观测数据变异性的能力,取值范围通常为0%~100%,越大的解释比例代表越好的拟合效果,即模型越能解释该区域的信号波动。另外,还可通过GLM模型[26]来评判解释性,如式(3)所示:
式中:β为系数向量;X为设计矩阵;e为误差向量。
将真实BOLD信号和经DCM拟合的信号代入式(3),分别反演这2种情况下的β,若两者的值具有一致的变化趋势,则说明DCM对所提取的BOLD信号具有较好的解释性。
-
DCM是将神经状态的变化描述为大脑各个区域间的因果连接活动、该连接受实验刺激的调制活动以及输入至大脑的刺激活动之和,其基本方程[14]为:
式中:z(t)为各脑区的神经状态;A为各脑区因果连接矩阵;B为输入调制矩阵;uj(t)为第j个输入刺激向量;U为刺激所构成的刺激矩阵;C为外部刺激输入的系数矩阵。该建模方程可用于揭示静态神经连接结构,也可动态地呈现大脑区域之间随着时间变化而产生的因果关系,还可以系统地揭示出心理学实验中不同条件变量对神经动态的调控模式。
式(4)中的DCM建模仍然在SPM12平台下完成,采用“DCM for fMRI”模块来进行参数估计。在DCM模型结构方面,参考面孔加工的经典神经通路,构建了一个rOFA与rFFA区域的双向连接模型,所有模型都设定这2个区域间同时存在前馈、反馈和自连接(即A矩阵的设计),并且允许rOFA与rFFA同时接收外部刺激驱动(即C矩阵的设计),以此模拟面孔视觉刺激的初始感知过程。
在刺激建模方面,包含输入刺激设计(即U的设计)及调制刺激设计(即uj的设计)。对于输入刺激设计,在本研究中未对不同面孔类型(如熟悉、陌生、打乱)或其重复类型(如立即重复、延迟重复)进行区分,而是统一视为一个广义“面孔呈现”的输入事件。与式(1)中所设计的回归因子一样,将式(4)中输入刺激所对应的回归因子向量U设计为:
该向量设计表示对所有呈现面孔的刺激均予以考虑,包括熟悉面孔、陌生面孔和打乱面孔的初始呈现、立即重复和延迟重复等,它可有效捕捉面孔加工通路对于视觉输入层面的整体激活趋势,对构建稳健的模型驱动项有一定帮助。该策略具有良好的生物学合理性,即初级输入在大脑皮层传播时一般不会因刺激类别产生明显变化,因此统一驱动输入更契合底层神经通路的实际机制[27]。与仅针对特定面孔类别(如陌生面孔)设置输入驱动的传统策略相比,该方法提供了更全面且统一的输入建模方式,保证建模过程中每类刺激都有相应的驱动贡献,提升了模型对BOLD信号的拟合能力与泛化能力[28]。
在调制刺激建模方面(即uj的设计),没有采用传统的二元对比策略(如将陌生面孔与重复面孔进行二值化处理)[29-30],而是引入多通道输入矩阵,即将实验中的不同刺激类型,比如“熟悉面孔-立即重复”“陌生面孔-延迟重复”“打乱面孔-立即重复”等,当作独立的输入维度和调制变量。该设计不仅可提高输入层建模的生物学合理性和心理学可解释性,还可从模型层面上剥离不同刺激条件间的潜在混淆效应。另外,该设计与组合对比[10]也不同,后者借助对不同实验条件进行线性组合来构造对比向量,但会导致调制变量之间存在重叠,容易引入调制混淆和变量共线性的问题。本研究独立地对每个实验条件的调制效应进行建模,提升了模型路径清晰度与参数解释可靠性,这符合减少变量共线性来提高B矩阵估计稳定性的原则[14]。同时,本研究更契合DCM建模假设,即特定心理变量调制特定神经连接,而不是多个复合条件共同起作用,因此能清晰识别和归因特定实验条件对具体连接产生的调制作用,避免传统联合对比设计下调制效应分布不明确等问题[28, 31]。同式(1)中所涉及的回归因子相同,将式(4)中的输入刺激向量uj(t)所对应的回归因子向量uj设计如下:
式中:u1-u6分别对应“熟悉面孔-立即重复”([0 1 0 0 0 0 0 0 0])、“熟悉面孔-延迟重复”([0 0 1 0 0 0 0 0 0])、“陌生面孔-立即重复”([0 0 0 0 1 0 0 0 0])、“陌生面孔-延迟重复”([0 0 0 0 0 1 0 0 0])、“打乱面孔-立即重复”([0 0 0 0 0 0 0 1 0])以及“打乱面孔-延迟重复”([0 0 0 0 0 0 0 0 1])。每个调制输入变量都明确对应一个实验条件,保障了调制效应的独立识别与清晰解释。
-
近年来,DCM与参数经验贝叶斯(Parametric Empirical Bayes,PEB)相结合的方法已成功应用于面孔识别任务中,有效揭示了不同刺激条件对关键脑区之间连接的调制作用[10]。该结合通过在群体水平上迭代更新个体模型参数,生成更合理的经验先验,从而提高了模型拟合的稳定性与生物学解释力[10, 32]。
PEB基于贝叶斯层级建模框架,采用超先验对个体参数的不确定性进行有效加权,使得组水平推断对样本量的依赖相对较小[28, 32]。研究表明,即使在小样本条件下,PEB仍能提供稳健且可靠的参数估计与贝叶斯模型证据推断[33]。此外,PEB结合贝叶斯模型简约和贝叶斯模型平均的方法,有效增强了贝叶斯因子和参数估计在有限样本下的稳定性[32, 34]。
本研究采用上述建模框架,先通过SPM12中的spm_dcm_peb_fit函数对每位被试的完整DCM模型进行估计,并在群体水平构建PEB模型。在该函数中,将M.X设置为ones(N,1),代表采用无协变量的一阶截距模型;M.Q设置为single,代表使用单一精度成分构建随机效应。另外,鉴于本研究重点关注因果模型中的连接调制,模型推断重点放在对B矩阵的推断,即评估不同类型刺激是否对rOFA与rFFA之间的连接通路(包括双向连接与自连接)产生显著调制作用。
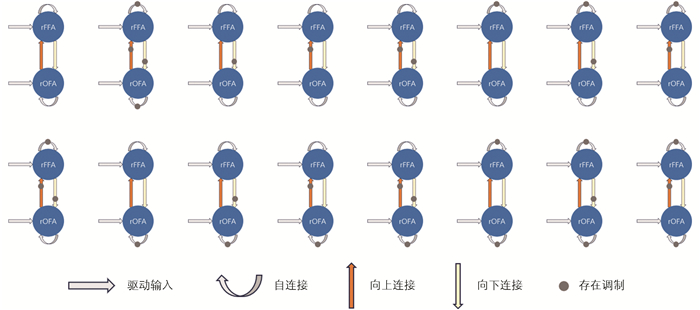
在候选模型空间中,通过4个不同的自连接和4个不同的互连接组合,系统构建了16个模型,其中每个模型均有不同的调制路径,如图 2所示。再通过6个调制通路家族来比较16个模型,评估每族连接是否在组水平上显著偏离零。该6个调制通路家族为:①任意自连接调制;②任意rOFA↔rFFA间调制;③ rOFA自连接调制;④ rFFA自连接调制;⑤ rOFA→rFFA前馈调制;⑥ rFFA→rOFA反馈调制。然后,计算6种调制刺激条件下各通路家族的第2种最优模型后验概率作为最终的模型后验概率,由其来检验连接是否被调制。第2种后验概率表示真实模型属于该家族的概率,相比第1种概率(对应于该家族不包含真实模型的概率),提供了该连接通路在群体水平下受到调制的量化证据,广泛应用于PEB的家族层面推断中[35]。上述的模型比较调用spm_dcm_peb_bmc函数完成,后验概率计算调用spm_dcm_peb_bmc_fam函数完成。
由于本部分调制路径理论上有明确方向预期,组级后验概率分析采用Pp>0.80作为调制效应的“趋势性证据”阈值[28]。当前DCM-PEB分析中,后验概率Pp>0.95被普遍视为强证据标准,用于判定连接在群体水平上的显著存在[28, 35]。然而,PEB属于贝叶斯概率推断框架,其本质是基于相对证据强度的概率模型选择。如Zeidman等[28]指出,低于0.95的概率亦可被视为“relatively strong probability”,在实证研究中具有明确的解释价值。尤其在样本量有限、信号质量受限或模型结构复杂的情况下,采用更低的概率阈值作为趋势性证据是合理且被接受的。因此,本研究将Pp≥0.95的路径视为“较大调制”,0.80<Pp<0.95的路径视为“可能存在调制趋势”,这种基于贝叶斯推断的不确定性表达方式提升了建模结果的解释弹性,也为理解面孔重复加工中的区域间因果交互提供了坚实的统计依据。
1.1. fMRI数据来源
1.2. 数据预处理与ROI提取
1.3. DCM结构与输入建模
1.4. 基于DCM的PEB推断
-
本节先给出“最大激活点覆盖”策略所确定的rOFA与rFFA中心坐标结果,如表 2所示,具体位置如图 3所示。在每位被试标准化空间中的fMRI数据上提取该球形区域内体素的第1个主成分作为BOLD信号,用于后续DCM建模分析。在采集过程中,本数据集中有5名被试在rOFA或rFFA区域未能获得有效的时序信号,表现为根据最优坐标构建的球形ROI掩模内体素数量为零。推测原因有个体解剖差异、标准化过程中的空间扭曲、头动导致的局部信号退化或目标区域缺乏有效激活。根据这一明确的空间有效性标准(即ROI掩模内体素数需大于零),为保障DCM模型结构拟合的准确性与参数估计的稳健性,本研究去除5名被试的数据,最终挑选了11名被试的数据。
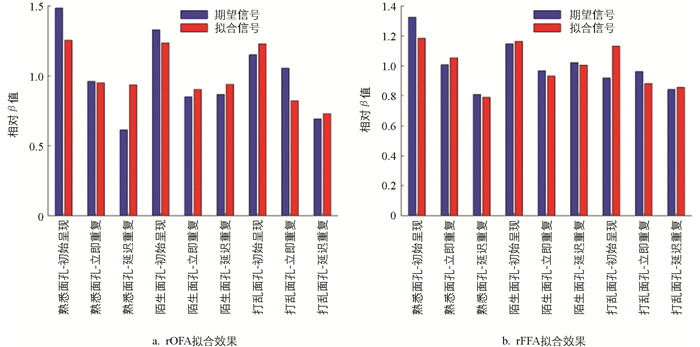
表 2还给出了rOFA和rFFA区域的拟合信号解释率,平均解释率分别为12.13%和10.39%,虽然略低于部分文献报告的区间16%~35%[10],但考虑到本研究中信号提取策略(基于球形感兴趣体积VOI且限制在激活峰附近)、被试样本规模(11个被试)及任务条件复杂度,该解释比例仍在DCM应用可接受的范围中,在捕捉区域性神经动态方面有一定合理性。为说明该合理性,对ROI半径进行了敏感性分析,在保持原始ROI中心不变的情况下,将球半径由10 mm扩大至12 mm,重新提取BOLD时间序列并拟合DCM。结果显示,rOFA的平均方差解释率变化极小(由12.13%增至12.17%),而rFFA的解释率反而降至-1.99%,出现拟合失败。这表明在本研究采用的基于激活峰的ROI框架下,简单扩大球半径并不能有效提升拟合性能,反而可能由于包含非目标功能区域的噪声信号,降低了信号质量。因此,当前的ROI半径设定在信号特异性与空间覆盖之间达到了较好的平衡。为进一步检验模型拟合的有效性,图 4给出了ROI提取信号与对应DCM拟合信号所反演的β值,该值为式(3)中向量β的幅度系数部分。从该结果中可以看到,对于所选定的rOFA和rFFA区域,模型拟合序列与所提取的真实序列的β值具有近似的变化趋势,因此表明拟合序列也可以重现真实序列的变化特征。
-
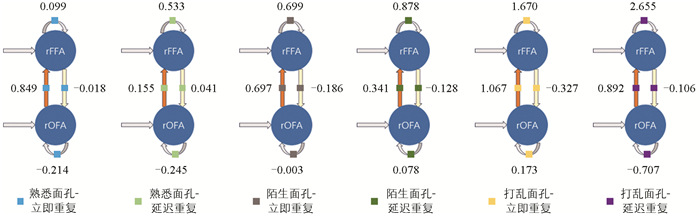
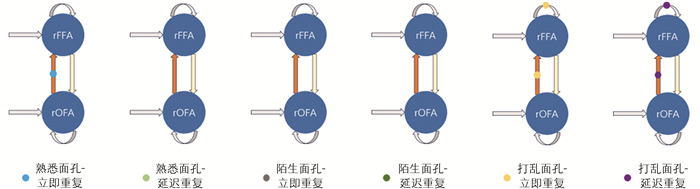
基于PEB和DCM的分析方法,本节对最终结果进行可视化,图 5给出了PEB推断的B矩阵参数的数值(即调制连接强度),表 3给出了刺激条件下各通路家族的后验概率,设置阈值后得到图 6的调制连接图。
结果显示,所有重复条件下反馈调制模型族(rFFA→rOFA)的后验概率都没有上升,即当前数据没有给出支持反馈路径调制的证据。这一结果表明,在本研究的实验结构与刺激设计下,自上向下的信息调制路径不是重复效应的主要承载机制,初步排除了“高阶预测驱动加工”主导的可能性(即高级视觉区域依据先前经验或者语义预期来生成预测信号,该信号借助反馈连接传递到初级区域,形成对即将到来的感知输入展开解释或者抑制加工的机制)。该理论一般是在预测编码框架下提出的,其观点为感知过程是大脑持续生成并修正针对外界输入的预测过程,但当前数据并未呈现出反馈通路的调制作用,这意味着基于高阶表征的预测性反馈在本实验的重复条件中未起到核心作用。
相对而言,rOFA→rFFA前馈调制模型族在多个条件下获得较大支持,在“熟悉面孔-立即重复”这一经典重复抑制范式中,该模型族的后验概率比所有竞争模型都高,说明重复加工的核心机制可能集中在初级面孔区域向高级表征区域的信息流动调制上。同时在该条件下,rOFA→rFFA的调制连接强度为0.849 Hz(需注意后验概率大并不意味调制强度大,前者表示调制通道存在的概率,后者表示调制通道的强度和方向),表明重复刺激增强了前馈连接强度,与“感知驱动”假设(即重复效应主要源于感觉通路对输入刺激的加工优化,并非高级区域生成预测后反馈到初级区域做调整)存在一定偏离。在该假设下,信息流是自下而上的,神经系统对重复刺激响应减弱体现出对物理输入的快速识别、效率提高以及神经资源的节省,而非认知预期的更新或者预测误差信号的抑制。而本研究观察到的正向调制却提示,在“熟悉面孔-立即重复”条件下,前馈路径或因经验表征的激活得到强化,这体现出系统对于语义相关输入的快速动员以及加工能力的提高,熟悉面孔的重复抑制机制并非完全是由感觉输入的节能型抑制产生的,而有可能涉及对语义信息的主动调配以及连接提高。本研究结果更偏向于支持一种将感知输入与经验调节相融合的重复机制解释,而不是仅仅依靠低阶感知优化或者高阶预测误差信号的被动抑制模型。
在“打乱面孔-立即重复”以及“打乱面孔-延迟重复”2种条件下,前馈调制模型族和rFFA自连接调制模型族呈现出较强的模型证据,这意味着即便面孔完整性遭到破坏,系统依旧可激活前馈通路,并且对rFFA内部的加工机制实施调制。在这2种条件下的rOFA→rFFA前馈调制连接强度均为正值(1.067 Hz和0.892 Hz),提示该重复刺激增强了初级区域向高级区域的信息传递;同时,rFFA的自连接调制强度也均为显著正值(1.670 Hz和2.655 Hz)。根据DCM中自连接参数的解释,正向自连接调制并不表示局部活动直接增强,而是表示rFFA区域的自抑制增强,提示该区域在处理打乱面孔重复输入时可能受到更强的局部抑制性调节。该结果或许体现出神经系统在面对结构不完整但重复出现的视觉输入时,一方面增强前馈信息传递,另一方面通过提高rFFA的局部自抑制,对不稳定或难以整合的表征进行约束与调节。
相对而言,在“熟悉面孔-延迟重复”“陌生面孔-立即/延迟重复”这些条件下,所有模型族的后验概率都没有呈现出明显优势。在这些条件下的重复刺激,未能在群体层面诱发出稳定的网络连接调制,这种结果可能与识别经验、面孔表征的稳定性,或者重复间隔对神经加工优先级的影响存在关联。
由图 5和图 6发现:①面孔重复的连接调制主要集中在前馈通路rOFA→rFFA上,在“熟悉面孔-立即重复”以及“打乱面孔-立即/延迟重复”条件下显现,且该前馈连接的调制强度均为正值,反映重复刺激增强了初级区域向高级区域的信息传递;②反馈连接的调制作用整体较为微弱,在所有重复条件中没有获得一致性证据;③自连接的调制作用有很强的条件依赖性,仅在打乱面孔的重复刺激下,rFFA的自连接显示出较强的正向调制,反映出该区域局部自抑制增强。综上,有效连接调制呈现出较大的单向性以及刺激条件的依赖性,重复加工过程受到时间结构、语义属性以及完整感知交互作用的限制。
为验证阈值设定的合理性,进一步分析了各连接后验概率的分布。结果显示,若采用Pp<0.70作为存在判定,将导致大部分连接被认定为存在,缺乏有效区分;而在0.70≤Pp≤0.80,新增的连接较少,且一致性明显低于Pp>0.80的连接。相较于Pp>0.95,Pp>0.80多出的2个连接,其概率为0.80~0.83,即在被试群体中有8~9人支持,仅2~3人不支持,表现出较高的组内一致性。相反,Pp=0.77的连接在个体间的一致性相对较低。因此,Pp>0.80在本研究中可以合理视为“趋势性证据”阈值,用于补充强证据之外的解释。
2.1. ROI序列
2.2. 有效连接分析
-
本研究运用DCM-PEB框架考察了面孔重复刺激下的调制加工通路,探究不同的语义属性以及不同时间结构的面孔重复刺激,以及在rOFA与rFFA之间的有效连接上是否呈现出各自独立且可区分的调制效应。为此,本研究采用了与传统组合对比所不同的独立对比方法:在ROI选取上,定位于rOFA、rFFA脑区中立即重复与延迟重复存在激活差异的区域;在调制刺激设计上,设置了6种独立的重复刺激(即“熟悉面孔-立即/延迟重复”“陌生面孔-立即/延迟重复”以及“打乱面孔-立即/延迟重复”)向量。
在输入刺激(DCM中C矩阵)设计上,本研究采用统一驱动输入,将所有面孔呈现作为一个共同的外部输入,未对不同面孔类别单独建模,主要基于2点考虑:首先,低级视觉区域在早期尚不体现类别选择性,类别加工主要出现在更高级的语义阶段[36],因此,统一输入既符合底层感知的共性特征[10, 28, 37],也简化了模型,避免自由度膨胀,同时提升稳健性;其次,DCM的驱动输入用于捕捉外部刺激进入神经网络的路径,实验条件差异则通过B矩阵的调制体现[38]。当然,也有研究采用类别特异的驱动输入[39-40],用于刻画不同刺激对下游区域的直接驱动,但会显著增加模型复杂度,尤其在类别与重复交叉时,易导致模型不稳定,贝叶斯证据下降[28, 41]。鉴于本研究关注的是立即与延迟重复如何通过连接调制作用于rOFA与rFFA,驱动输入的类别划分并非核心目标,因此单一驱动输入的劣势(即无法捕捉类别间潜在的直接驱动差异)在本研究中也就不存在。通过B矩阵对重复条件和面孔类型建模,能够有效反映实验关心的神经机制。因此,统一驱动输入不仅具备神经生理合理性,也符合DCM的简约性原则[42-43]。近年来,多项研究亦倾向采用此策略,借助有效连接的调制反映实验条件差异,更契合神经加工的层次结构,使模型更直接捕捉刺激对连接动态的影响[10, 42, 44]。本研究沿用这一方法,以揭示重复刺激如何通过调节rOFA与rFFA的连接实现功能适应。
研究结果表明,重复相关调制主要集中在rOFA→rFFA的前馈连接上,尤其体现在“熟悉面孔-立即重复”以及打乱面孔的重复条件下。反馈连接在所有重复条件中均未获得稳定证据,而自连接调制则具有明显的条件依赖性,仅在打乱面孔重复条件下表现为rFFA自连接的正向调制。
需要指出的是,在DCM框架下,调制连接的正负并不直接对应于BOLD信号的升高或降低。传统fMRI中,重复抑制表现为特定脑区BOLD信号下降,反映出局部神经活动的整体减少,而DCM描述的是神经状态变化率之间的因果耦合,BOLD信号则是神经状态经血流动力学卷积后的观测结果。因此,前馈连接的增强(如rOFA→rFFA)意味着源区对靶区的驱动效应在重复条件下增强,并不与BOLD信号的减弱矛盾,反而可能反映出信息传递的效率提升,即通过强化前馈驱动,系统能更快速高效地处理重复刺激,减少对冗余神经资源的依赖,从而在BOLD信号上表现为下降。
从预测编码的视角[45],重复刺激提升了内部模型的预测精度,降低了对预测误差的依赖。前馈连接增强体现了精简误差信号的高效传递,而BOLD信号下降反映了误差减少。已有研究指出,RS不仅表现为神经反应振幅降低,还伴随更高效的信息加工机制[46]。从建模上看,DCM中的调制连接(B矩阵)调节的是状态变化率的耦合强度(如rOFA→rFFA增强表示rOFA状态变化对rFFA的影响更强),是否引起BOLD信号升高,取决于前馈、自连接(A矩阵)和外部输入(C矩阵)等因素的共同作用。BOLD反映的是系统状态随时间的积分,因此前馈连接增强与BOLD信号下降在数学上完全共存。前馈连接增强与BOLD层面的重复抑制并不矛盾,而是共同反映了神经系统在信息传递效率与能量消耗之间的优化平衡。
本研究仅发现了向上传递方向的调制效应,这一结果与Ewbank等[17]在面孔尺寸保持不变的条件下观察到“仅存在向上RS”的结果十分吻合,提示该效应可能受到输入路径结构以及刺激属性的共同约束,推测这种连接模式的形成可能和实验范式中面孔信息同步进入rOFA与rFFA的处理路径有关。已有研究说明,rOFA对面孔的初级物理特征(如尺寸与方位)具有更高的敏感性[47],而rFFA更侧重于整合全局特征与语义信息,在当前没有尺寸或视角变化的条件下,信息流动缺乏十分突出的处理层次差异,没有激活典型的由高级区域向低级区域调制的需求,这解释了为何本研究在所有重复条件下都没有观察到反馈方向的连接调制现象。
进一步看,本研究采用的实验范式属于隐式任务,参与者无需对面孔的身份、情绪或其他语义信息进行显式判断。这种范式倾向于主要驱动底层视觉加工通路,反馈通路的激活需求较低,这一点在最近的研究中也得到了验证。Garlichs等[29]在一项显式面孔识别任务中发现,面孔识别过程中存在显著的反馈调制,尤其是在OFA中表现出反馈驱动的特征锐化现象。与此相反,本研究采用的隐式任务未能观察到类似的反馈效应,这一差异表明,反馈通路的激活在很大程度上依赖于任务对高阶语义信息加工的需求。显式任务强调身份判别、语义推断等高阶认知操作,从而激活自FFA乃至更高级皮层向OFA的反向调制,以优化早期视觉表征。而在隐式范式下,参与者并不需要对面孔语义进行主动处理,因此反馈通路的调制动力不足,连接模式主要表现为前馈增强。
此外,从预测编码的角度来看,反馈连接通常承担着传递高层预测的作用,用以约束下级区域的感知输入[45]。在显式任务中,高阶认知状态(如面孔身份的先验预测)被充分激活,驱动更强的反馈调制;而隐式任务缺乏这种明确的预测目标,系统更多依赖于前馈误差信号的上传,从而形成本研究观察到的以前馈为主导的连接模式。
-
在“熟悉面孔-立即重复”条件下,rOFA→rFFA的前馈连接呈现出正向调制情况,这意味着信息在自下而上进行传递期间获得了增强,表明即便面孔为熟悉,但当神经系统重复接收到该面孔呈现时,依然会强化初级区域至高级区域之间的信息传递过程,这可能意味着对识别结果进行确认性加工,或是激活与之相关的语义信息,这种加工模式与传统重复抑制理论中“输入节省”的阐释并不完全契合,反而更有可能呈现出一种以记忆驱动为核心的加工提高机制,即熟悉面孔的重复出现涉及知觉匹配,还可能激活与个体经验相关联的语义以及身份表征。与之相比,“熟悉面孔-延迟重复”并没有引发类似或相反的连接调制现象,这可能是因为重复间隔的增加使得刺激之间的表征匹配效应被削弱了,其与重复启动效应变弱的现象一致,即随着时间间隔不断延长,神经响应的节省效应(如BOLD减弱或者连接抑制)不再明显[24]。
-
对于陌生面孔的重复,在任何一种时间结构下,都未曾引发有效连接的调制变化,这样的结果或许反映出了任务加工深度同记忆系统激活之间存在的互动关联。在本研究采用的隐式任务中,也即判断面孔对称性而非识别身份,被试并没有主动去调动与面孔相关的语义或者记忆路径,使得表征更新以及预测机制没能被有效激活。
有研究[48-49]显示,熟悉刺激能更明显地激活涉及长期记忆以及情绪加工的脑区(如海马、杏仁核),而陌生面孔在无意识识别的背景之下,大多缺少与语义属性的匹配过程,很难产生典型的RS效应[18, 20]。为了进一步验证这一神经机制,本研究基于现有fMRI数据进行了全脑对比分析(“熟悉面孔-立即重复”大于“陌生面孔-立即重复”)。结果显示,在MNI坐标为(18,-34,-16)的右侧海马存在显著激活(p<0.001,无校正)。该结果表明,熟悉面孔的重复能够有效调动海马相关的长期记忆通路,而陌生面孔则缺乏这一加工路径的参与。这一发现为本研究在有效连接层面未观察到陌生面孔的调制效应提供了直接的神经支持。
从功能机制上看,海马与内侧颞叶系统在面孔表征更新中的作用,主要体现在将感知输入与既有的身份表征相匹配,从而驱动预测模型的形成和误差信号的修正[50-51]。在缺乏这种身份-语义联结的陌生面孔条件下,记忆-语义网络无法有效介入,导致系统仅依赖底层视觉路径的被动加工,难以形成有效的表征更新机制。尽管本研究的DCM模型主要聚焦于视觉通路(rOFA和rFFA),但缺乏来自内侧颞叶、杏仁核或前额叶的上行调制,可能导致系统在面对陌生面孔重复时,无法形成有效的预测模型或表征更新机制。
另外Stam等[52]也指出,RS不应该仅仅被看作是预测误差最小化的体现,其本质可能更多地受到初始刺激强度以及刺激熟悉度调制方面的影响。在连接层面没有观察到的对于陌生面孔的重复调制也支持这一观点,并且对将RS单纯归因于预测编码的传统理论提出了修正性的补充。
-
打乱面孔的重复加工呈现出另外一种明显不同的连接特性,本研究观察到它引发了rOFA→rFFA前馈连接的正向调制,并且在rFFA的自连接方面也呈现出正向调制,即使面孔的结构完整性遭到破坏,其局部特征依然可引发神经系统的特异性加工反应。已有研究说明,OFA主要处理面孔的局部部件信息,而FFA负责整合这些部件以构建整体面孔表征[53]。在打乱结构的情况下,尽管整合难度增加,但局部特征的重复或许仍对自下而上的传递起到了强化作用,呈现出rOFA→rFFA的连接增强。rFFA可能通过更强的局部抑制性调节,对难以形成稳定整体表征的输入进行约束,从而避免非典型或不稳定表征在高级面孔区域中被持续放大。
-
从机制方面看,熟悉面孔的重复效应更多体现了长期记忆参与下的语义驱动加工路径,打乱面孔的增强效应更接近感知驱动的局部匹配机制,而陌生面孔在隐式任务里没能激活语义网络,缺乏明确的连接调制模式。本研究推测,面孔RS效应可能反映出一种更为灵活的连接调制趋势,未来的研究可在更为细化的任务范式里,剖析长期记忆与语义属性的贡献比例,或者借助操纵语义属性以及隐式任务的注意水平,去验证感知驱动与长期记忆在RS调制中的功能边界。另外,可考虑把该模型应用到其他相关的重复刺激里,比如姓名、声音或者情绪面孔等,以此拓展对于重复的时间结构、语义属性以及完整感知等多重因素调控的跨模态理解。
-
本研究依据DCM-PEB框架系统地指出面孔重复条件下神经通路中有效连接的调制模式,此调制并非统一或者自动触发的机制,而是高度依赖于重复时间结构、语义属性与完整感知之间的复杂交互。实验发现,在“熟悉面孔-立即重复”条件中,rOFA→rFFA的前馈连接呈现出正向调制,这揭示了经验性表征在塑造连接调控过程中发挥着核心作用。相比之下,陌生面孔在所有重复条件下都没有引发连接水平的较大调制,而打乱面孔尽管缺少身份语义信息,却在不同重复时间结构下调制了前馈通路与自连接,这暗示结构相似性也可驱动神经加工路径的功能重构。上述结果共同支持以下结论:
1) 重复抑制体现的是一种“条件感知的连接重编程”机制,即神经系统在面对重复输入时,会依据刺激的重复时间结构、语义属性以及完整感知的适配关系,选择性地调制特定方向的连接路径。这一机制突破了重复加工被动抑制的传统假设,更强调神经网络具备主动、条件响应的可塑性特征。
2) 本研究所采用的隐式任务范式揭示,即使在非显式判断的情境中,不同重复类型已能在连接层面呈现出功能性分化的调制模式,这进一步佐证了神经系统对重复信息的加工策略具备条件驱动的可变性,而非一套固定通路。
以上结论为理解面孔识别等复杂语义加工中的连接机制提供了新的证据,为检验时间结构、语义属性以及完整感知的交互对神经连接调制的普适性和特异性提供了理论基础。