


 下载:
下载:
-
大量水生生物(如浮游生物、藻类、溪流昆虫等)生活在河流、溪流等具有单向水流的生态系统中. 为何生物种群在受到从上游向下游单向水流不断冲刷的情形下能够生存?这是河流生态理论中一个重要的被称为“漂移悖论”(driftparadox)的科学问题. 为了解释该科学问题,多位科学工作者建立并研究了一系列包括对流和扩散过程的反应-扩散-对流方程模型和积分-微分方程模型[1-7]. 这些模型描述了河流种群的时空演变过程,其长期动力学行为(持久性或灭绝)通过多种数学理论和工具被深入研究,例如一致持久性、零解的稳定(不稳定)性、正稳态解的存在性和稳定性、临界区域长度、行波解和渐进传播速度等. 基于上诉文献中的单个种群模型,许多研究者建立了两个相互作用的河流种群模型,着重研究了水流的速度、种群的扩散系数等因素如何影响种群的共存(排斥),例如:文献[8-10]研究了两个竞争种群的模型;文献[11]研究了河流生态系统中的捕食-食饵种群的模型. 这些模型具有以下一般的形式:
其中:u(x,t),v(x,t)表示位置在x处、时间在t处两物种的密度;di(i=1,2)是扩散常数,qi(i=1,2)是对流系数(即水流速度),f(u,v)与g(u,v)为反应函数.
当q1=q2=0时,系统(1)变成经典的两个相互作用种群的反应-扩散方程模型. 反应-扩散方程模型中由扩散导致的常数稳态解的不稳定性称为Turing不稳定性.
本节的主要目的是研究反应-扩散-对流模型(1)中的单向水流(即对流项)对斑图动力学的影响(q1=q2=q). 为此,我们考虑如下长度为L的河流中两个相互作用种群的动力学模型:
其中的Neuman边界条件表示河流上游有随着河流自然流入的种群,河流下游随着时间流逝有被河水带走的种群.
全文HTML
-
本节讨论没有水流的影响,及q=0的情形下,系统(2)的Turing不稳定性.
假设(u*,v*)是系统(2)的常数稳态解,将系统(2)在(u*,v*)处线性化,令ϕ=u-u*,φ=v-v*,则有
其中f1=fu(u*,v*),f2=fv(u*,v*),g1=gu(u*,v*),g2=gv(u*,v*). 用矩阵表示为
其中
在一维Neuman边界条件情形下,系统(4)解的形式为
其中
$k=\frac{n \pi}{L}$ ,$n \in \mathbb{N}^{+}$ . 要寻求系统(4)的非平凡解,则需其中
如果系统(4)是一个线性不稳定的系统,那对于某些
$n \in \mathbb{N}^{+}$ ,当t ∞时,Ψ(x,t)将会趋于无穷. 即在(5)式的解中某个ω存在正实部,或者矩阵Mk=-k2D + J的某个特征值有正实部,这取决于Mk的迹和行列式值的符号:在(5)式中ω=ω(k)是决定稳态(u*,v*)稳定性的特征值. 对于k=0,(6)式的两个根满足
假设
这意味着稳态(u*,v*)在没有空间作用的情况下是线性稳定的. 现在对于k>0,(6)式的两个根满足
因为d1>0,d2>0并且假设(10)式成立,那对于所有
$k=\frac{n \pi}{L}$ ,$n \in \mathbb{N}^{+}$ 都有ω1k+ω2k < 0. 因此,如果对于所有k>0都有ω1kω2k>0,那么(u*,v*)仍然保持稳定. 如果对于某些k>0有ω1kω2k < 0,那么(u*,v*)变得不稳定,这种由扩散导致的不稳定性通常被称作Turing不稳定,意味着模式的形成.假设(10)式成立,当d1g2+d2f1>0时,Det(Mk)在k2=kmin2=
$\frac{d_1 g_2+d_2 f_1}{2 d_1 d_2}$ 处取得最小值. 这时Det(Mk)>0等价于(d1g2+d2f1)2-4d1d2|J|>0. 那么系统(2)的空间齐次稳态解(u*,v*)可能失去稳态当且仅当和
同时成立. 在(12),(13)式同时成立的情况下,将Det(Mk)=0的两根设为k12,k22
当存在
$n \in \mathbb{N}^{+}$ ,使得k2∈(k12,k22),那空间齐次稳态(u*,v*)就会失去稳定,发生Turing不稳定,就有空间模式的形成. 故模式的形成只需不等式k2-k1>$\frac{\pi}{L}$ 成立即可.经计算(k2-k1)2=
$\frac{\left(d_1 g_2+d_2 f_1\right)-2 \sqrt{d_1 d_1 \mid \boldsymbol{J}\mid}}{d_1 d_2}$ ,则k2-k1>$\frac{\pi}{L}$ 等价于事实上不等式(15)是不等式(13)的充分条件. 通过上面讨论得出如下定理.
定理1 假设(10),(12),(15)式同时成立的情况下,系统(2)的空间齐次稳态解(u*,v*)会发生Turing不稳定.
-
接下来考虑在发生Turing不稳定后,加入对流的情况,即系统(2)中q>0.
在一维的Neuman边界条件下,经计算可得dFxx-qFx=-λF的特征值分别为:λ0=0,对应特征函数
$F_0=C \mathrm{e}^{\frac{q}{d} x}$ ;λk=dk2+$\frac{q^2}{4 d}$ ,k=$\frac{n \pi}{L}$ ,$n \in \mathbb{N}^{+}$ ,对应特征函数Fk=C$\left[\mathrm{e}^{\frac{q}{2 d} x}\left(\frac{q}{2 k d} \sin k x+\cos k x\right)\right]$ .这时系统(4)解的形式变为
同上讨论,(6)式变成:
其中
Mk的迹和行列式值为:
已知ω1k+ω2k < 0.Det(Mk)=0可写成:
其中a=
$\frac{1}{16 d_1 d_2}$ ,b=$\left(\frac{d_1 k^2}{4 d_2}+\frac{d_2 k^2}{4 d_1}-\frac{g_2}{4 d_1}-\frac{f_1}{4 d_2}\right)$ ,c=d1d2k4-(d1g2+d2f1)k2+|J|.在q=0时,若系统(2)发生Turing不稳定则k2∈(k12,k22),此时d1d2k4-k2(d1g2+d2f1)+|J| < 0. 由此可知当q>0时,只有一个正解q*2=Q*,
此时若
$q < \sqrt{Q_*}$ 则ω1k+ω2k < 0,ω1kω2k < 0,空间齐次稳态解(u*,v*)仍然不稳定;若$q < \sqrt{Q_*}$ ω1k+ ω2k < 0,ω1kω2k>0,空间齐次稳态解(u*,v*)重新稳定. 综上讨论,得出以下结论:定理2 假设(10),(12),(15)式同时成立,系统(2)的空间齐次稳态解(u*,v*)会产生Turing不稳定. 加入对流项之后,当
$q < \sqrt{Q_*}$ 时,空间齐次稳态解仍然不稳定;当$q>\sqrt{Q_*}$ 时,空间齐次稳态解会重新稳定.
-
标准的Lotka-Volterra捕食-食饵模型假设单位捕食率只依赖于食饵的数量. 但在大多数情况下,特别是当捕食者需要花更多时间去寻找食物时,更适合的捕食-食饵系统应建立在所谓的比例依赖性理论上[12]. 也就是说,单位捕食者增长率应该是食饵与捕食者比值的函数. 文献[13-14]讨论了扩散驱动捕食-食饵模型产生空间结构的一些条件. 近年来一些研究者研究了比例依赖(即反应项具有收获率)的捕食-食饵模型[15-16]. 我们考虑生活在河流生态系统中的捕食和食饵种群的相互作用,研究以下具有捕食者收获率的反应-扩散-对流比例依赖捕食-食饵模型的时空模型
其中:u(x,t),u(v,t)分别代表食饵与捕食者的密度,r表示食饵的内在增长率,K代表环境容纳量,c代表捕获率,c1代表转化系数,m代表半捕获饱和常数,e代表捕食者死亡率,h代表收获率. 为了方便,对系统(19)进行无量纲化:
仍记变量为u,v,x,t,q,则系统(19)可以写成
其中:
$\alpha=\frac{c K}{m r}$ ,$\gamma=\frac{e}{r}$ ,$\beta=\frac{c_1 c}{r}$ ,$\delta=\frac{h}{r}$ ,$d=\frac{D_2}{D_1}$ .系统(20)对应的常微分系统为
计算可知模型(21)有唯一的正平衡点E*=(u*,v*)=
$\left(1-\alpha+\frac{\alpha(\gamma+\delta)}{\beta}, \left(\frac{\beta-\delta-\gamma}{\gamma+\delta}\right) u_*\right)$ 的充要条件是(H1) δ1 < δ < β-γ,其中δ1=max
$\left\{0, \beta-\gamma-\frac{\beta}{\alpha}\right\}$ .模型(21)在E*的Jacobian矩阵为
其中:
对应的特征多项式为
对于Jacobian矩阵J(u*,v*)有如下结论.
引理1 1) 如果(H1)成立,则f1g2-f2g1>0.
2) 如果
(H2)αβ2+β(γ+δ)2 < α(γ+δ)2+(γ+δ+1)β2成立,则f1+g2 < 0.
3) 如果
(H3)αβ2+β(γ+δ)2>α(γ+δ)2+(γ+δ+1)β2成立,则f1+g2>0.
通过对Jacobian矩阵J的讨论,可得出如下结论.
定理3 假设(H1)成立. 如果(H2)成立,系统(21)的唯一正平衡点E*是局部渐进稳定的;但如果(H3)成立,系统(21)的唯一正平衡点E*是不稳定的.
系统(21)正平衡点E*也是系统(20)的空间齐次稳态解. 由定理1知,发生Turing不稳定当且仅当
在发生Turing不稳定后, 根据定理2知, 当
$q>\sqrt{Q_*}$ , 不稳定的E* = (u*, v*) 空间齐次稳态解会重新稳定, 其中Q*的表达式见定理2.下面利用Matlab对上诉模型进行数值模拟,以此来验证以上理论分析的结果.
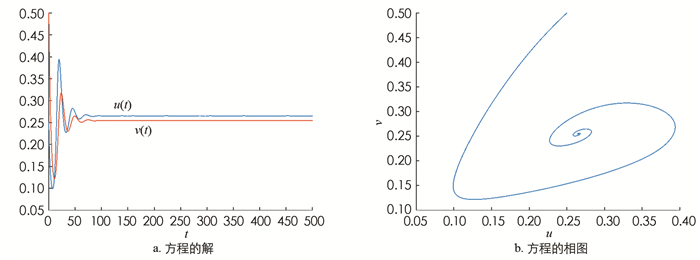
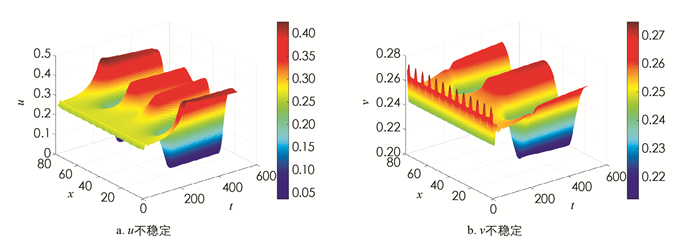
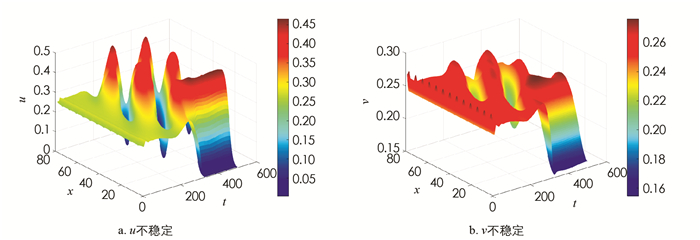
取参数为α=1.5,β=1.0,γ=0.01,δ=0.5. 此时正常数平衡点E*=(0.265,0.255)是渐近稳定的(图 1). 现加入扩散,取d=50,L=25π,(u0,v0)=(0.265+0.01sin(x),0.255+0.01sin(x)),(22)式的4个不等式均满足,此时发生Turing不稳定,有空间模式形成(图 2). 在发生不稳定后加入对流项. 在前面取定的参数情况下,计算得k12=0.121 2,k22=0.300 2,取n=4,则k2=0.16∈(k12,k22). 此时Q*=0.360 6,取q=0.2,
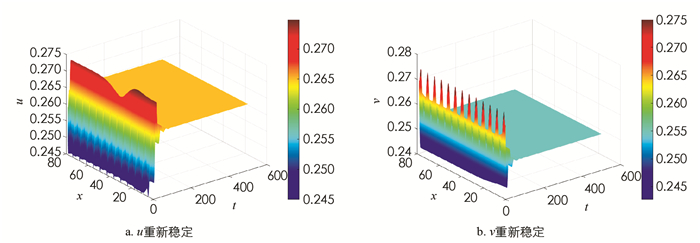
$q < \sqrt{Q_*}$ ,此时正常数平衡点仍然不稳定(图 3);但取q=1.5,则$q>\sqrt{Q_*}$ ,正常数平衡点重新稳定(图 4).
-
我们利用反应―扩散―对流方程来描述河流生态系统中的捕食者与食饵的相互作用,研究了在河流长度一定的条件下发生Turing不稳定性的一些必要条件,还研究了对流项对Turing不稳定的影响. 发现存在一个对流系数临界值,当对流系数大于临界值时会使发生Turing不稳定的常数平衡解重新稳定,但当对流系数小于临界值时,加入对流项对Turing不稳定性不产生影响. 但本文研究的情况比较特殊,本文中的边界条件是一维Neuman边界条件,还假设了对流项系数相等. 如果对流项系数不等,那么河流长度无限长等情况都有待进一步研究.