


 下载:
下载:


-
开放科学(资源服务)标识码(OSID):

-
干旱胁迫是全球农业生产力的严重制约因素,对农业生产构成持续威胁,干旱的强度和严重程度预计在不久的将来会持续增加. 农业生产中日益严重的缺水和频繁的干旱已经在世界范围内造成巨大的灾害,导致许多作物减产[1]. 大豆(Glycine max)是重要的油料作物、经济作物及饲料作物,在我国国民经济中占有重要地位. 然而大豆是一个对水分亏缺敏感的作物,在干旱胁迫下,大豆的产量损失可能高达40%,而长期或严重的干旱胁迫甚至会完全破坏种子生产[2-4]. 因此筛选挖掘大豆抗旱相关基因,解析其在大豆抗/耐旱中的功能和调控网络,将为培育抗/耐旱大豆新种质或新品种提供理论依据和有效基因,具有重要的科学意义和应用价值.
植物抵抗非生物胁迫是一个复杂的协调过程,涉及许多转录调控因子. 在这些转录调节因子中,G-box结合因子(G-box Binding Factor,简称GBF)能够通过识别靶基因启动子中的顺式作用调节元件G-box(CACGTG或CANNTG),进而调节靶基因的表达[5]. 已有研究表明G-box结合蛋白参与了多种细胞过程,在植物生长发育和抗/耐生物和非生物逆境方面具有重要作用[6-8]. B12D是一个G-box结合蛋白,该基因家族在植物发育和应激反应的各个方面中对细胞运输和代谢的调节起着重要作用[8-11]. B12D家族包含多个成员,B12Dg是该家族成员之一,最早在筛选大麦糊粉层和胚胎的转录组中发现,该基因启动子中含有赤霉素识别元件,在大麦种子发芽时期优势表达[8-9, 12]. 红薯的同源基因在衰老叶片中表达上调[13]. 拟南芥的B12Dg在种子萌发、植株生长、开花和荚果发育中均有表达[14]. 地黄(Rehmannia glutinosa)的B12Dg基因受铬、GA和盐胁迫诱导表达[11]. 小麦B12Dg在干旱胁迫下下调[15]. 甜菜(Beta vulgaris)B12Dg能够响应盐胁迫[16]. 多种基因型水稻,在水淹胁迫条件下都能检测到B12Dg基因的诱导表达,而且水淹诱导的基因启动子序列中含有G-box位点[17]. Almutairi[18]分析了14个物种中66个B12D蛋白,在预测的B12D启动子中发现了对光、脱落酸、茉莉酸甲酯、细胞分裂素、干旱和热响应的顺式调控元件. 这些研究说明GBF基因在植物响应非生物胁迫中具有重要作用.
为了筛选大豆抗/耐旱相关基因,本研究对大豆幼苗进行了干旱处理并对处理样品进行了转录组分析. Glyma.08G250300基因在干旱处理样品中显著上调,是一个干旱诱导基因(命名为GmDIBL1,Drought Induced B12D Like 1),可能在大豆抗/耐干旱胁迫中具有作用. 本研究进一步克隆了GmDIBL1基因的基因组序列和互补DNA(complementary DNA,cDNA)序列,分析了其序列特征,检测了该基因在大豆不同组织器官以及不同处理下的表达特性,进而通过构建过表达和基因编辑载体并进行大豆毛状根遗传转化,鉴定了GmDIBL1在大豆耐旱中的功能,以期为深入研究GmDIBL1在大豆抗/耐旱中的生物功能和调控机制奠定基础.
全文HTML
-
试验用大豆品种为‘Williams 82’ ‘晋豆21’和‘绥农26’. 使用的菌株有大肠杆菌菌株感受态细胞DH5α、根癌农杆菌感受态GV3101和发根农杆菌感受态K599.
-
挑选大小和活力一致的‘晋豆21’和‘绥农26’种子于培养箱萌发、生长,生长环境为温度25 ℃,空气相对湿度60%~70%,长日照(昼16 h/夜8 h). 待幼苗的第一对真叶展开后,将大豆幼苗从土壤中轻轻拔出根部,用自来水冲洗根部表面泥土,再用滤纸吸干根部表面水分,用于干旱处理. 干旱处理首先将大豆根部的营养土清洗干净,置于15% PEG6000的1/2MS营养液中进行干旱胁迫. 分别在处理后0 h、6 h、12 h的时间点取植株根样品,做好样品编号. 每个取样时间点取3个生物学重复. 所有样品均用液氮快速冷冻,并在-80 ℃保存备用.
-
挑选大小和活力一致的‘Williams 82’大豆种子于自然条件下萌发、生长,取其根、茎、叶、花、根瘤、荚样品,取3个生物学重复. 使用Trizol方法,提取大豆不同组织以及干旱处理样品的总RNA,并进行琼脂糖凝胶电泳检测.
-
提取得到的RNA样品按照反转录试剂盒(TaKaRa,北京)说明书的具体操作步骤,合成大豆cDNA,-20 ℃保存. 以大豆根cDNA为模板,DIBL1-F/DIBL1-R为引物(DIBL1-F:5'-ttggagaggacagggtaccATGGGGCGTTGGATGAAACCA-3',DIBL1-R:5'-gccatggatccccgggtaccTTTATCCTCGCTGAAGAAGT-3'),聚合酶链式反应(Polymerase Chain Reaction,简称PCR)扩增得到GmDIBL1.
-
以大豆GmDIBL1编码的蛋白为检索序列,利用TBtools软件进行序列同源性比对和相似性搜索. 根据InterPro数据库(
https://www.ebi.ac.uk/interpro/result/InterProScan/ )预测GmDIBL1编码蛋白的结构域. 根据Panther数据库分析GmDIBL1编码蛋白的蛋白质家族类型. 利用跨膜结构域分析(TMHMM)预测GmDIBL1编码蛋白的跨膜结构域. 根据Phobius预测GmDIBL1编码蛋白的蛋白结构. 选择与大豆GmDIBL1编码蛋白的高度同源序列,利用MEGA11构建进化树. -
运用实时荧光定量聚合酶链式反应(Real-time PCR,简称RT-PCR)扩增分析GmDIBL1在不同组织以及干旱处理下的表达水平. 用大豆Actin作内参基因,Actin引物为GmActin-F(5'-CCGGTCGTGACCTCACTGATTTCT-3')和GmActin-R(5'-CATCAGGCAACTCGTAGCTCTTCT-3'),GmDIBL1的引物为GmDIBL1-qRT-F(5'-GGGCGTTGGATGAAACCAGA-3')和GmDIBL1-qRT-R(5'-TCTCTGGTGGTCGAGTACGA-3').
-
选择限制性内切酶KpnⅠ对载体pCAMBIA2300-35s-dsred-eGFP进行酶切,并对酶切后的产物进行胶回收,将GmDIBL1基因片段的回收产物与酶切线性化pCAMBIA2300-35s-dsred-eGFP载体片段的回收产物进行重组反应.
-
以pTX2262为模板,KO-DIBL1-F/KO-DIBL1-R为引物,通过PCR得到双靶点目的片段,并使用Golden Gate连接方式与骨架载体pTX2262连接.
-
将不含终止密码子的编码序列(Coding sequence,简称CDS)片段插入pCAMBIA2300-35S-GFP载体,获得重组质粒35S∷GmDIBL1∷GFP并导入根癌农杆菌GV3101. 将含有重组质粒的农杆菌菌液注射到生长旺盛的本氏烟草叶片中,暗培养过夜后正常光照培养1 d,使用DAPI染料对烟草叶片染色10~20 min,使用激光共聚焦显微镜观察烟草叶片细胞中的荧光信号.
-
将大豆种子种入土中大约生长1周龄左右,对大豆幼苗进行农杆菌侵染. 在大豆种子萌发生长期间准备农杆菌材料,将构建好的过表达载体和基因编辑载体以及对应空载体转入发根农杆菌K599的感受态细胞中,并将其涂于含有卡那霉素和链霉素的固体培养基中让其生长1~2 d. 用枪头挑取在上述培养基中生长的农杆菌菌落并活化1 d,吸取活化后的菌液300 μL涂在上述培养基上,于28 ℃恒温培养箱中培养1~2 d后可长成菌膜,平板上的菌膜用刀片刮成菌堆. 将大豆幼苗在子叶下1 cm处沿45°斜切,切口从菌堆中蘸取转入不同质粒的K599菌,将蘸取K599菌的大豆幼苗直接插入蛭石中,放入培养箱中培养10~14 d,培养条件为温度26 ℃,空气相对湿度60%,长日照(昼16 h/夜8 h).
-
两周后,取出植株,在双波长激发光源下检测阳性根,每个植株只留1条健壮的阳性根,其他阳性根和阴性根剔除干净. 同时提取过表达毛状根的RNA进行RT-PCR以确保该基因在根中过量表达,提取基因编辑毛状根的DNA并测序验证以确保该基因被敲除. 使用15%PEG6000处理大豆毛根转化复合体,随时观察其在15% PEG6000处理下的表型. 干旱处理24 h后拍照收集大豆植株表型.
1.1. 植物材料与菌株
1.2. 大豆种植及模拟干旱处理
1.3. 植物RNA的提取
1.4. 大豆GmDIBL1的扩增
1.5. 基因的生物信息学分析
1.6. 基因表达分析
1.7. 过表达载体的构建
1.8. 基因编辑载体的构建
1.9. 亚细胞定位检测
1.10. 大豆毛根转化复合体植株的产生
1.11. GmDIBL1大豆毛根转化复合体植株的表型鉴定
-
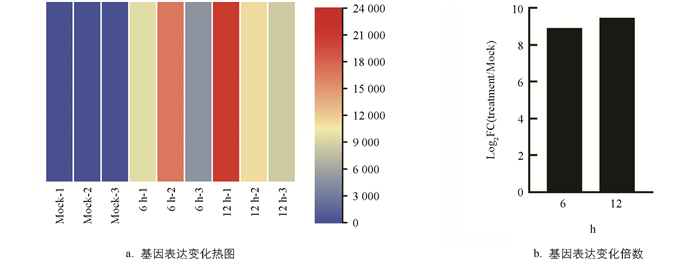
为了筛选大豆的干旱胁迫相关基因,利用PEG6000处理大豆‘绥农26’幼苗,分别取处理0 h、6 h和12 h的根进行转录组测序分析. 结果表明,GmDIBL1(Drought Induced B12D Like 1)基因在干旱处理的根中表达水平显著增加(图 1a),该基因在干旱处理6 h和12 h的根中表达水平分别增加了477倍和705倍(图 1b). 这一结果说明GmDIBL1基因是一个受干旱诱导的基因,可能在大豆响应干旱胁迫中具有重要作用.
-
为了获得GmDIBL1基因的序列,以公共数据库(
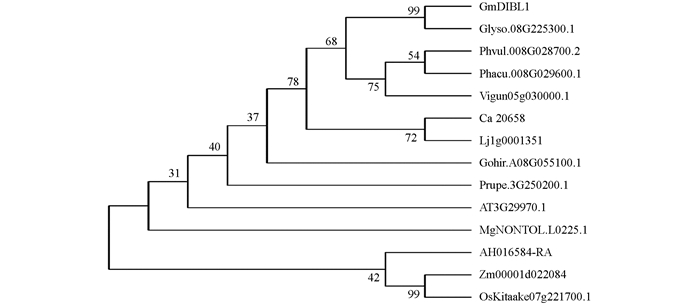
https://www.soybase.org/ )中Glyma.08G250300基因的序列为参考序列,设计扩增该基因CDS的引物. 进而以大豆‘williams 82’的根cDNA为模板进行扩增,结果获得长261 bp的cDNA序列,该序列编码86个氨基酸残基(图 2),推导的分子量为10.1 kD,等电点为9.647. 根据InterPro数据库(https://www.ebi.ac.uk/interpro/result/InterProScan/ )预测,结果显示GmDIBL1的蛋白质序列包含B12D结构域(IPR010530和PF06522,从氨基酸残基6-73)(图 2a),根据Panther分析,GmDIBL1是G-box结合蛋白(PTHR33417). TMHMM分析显示该蛋白质序列中含有一个跨膜结构域(氨基酸残基10~32),GmDIBL1含有信号肽序列(1~33 AA),包含信号肽N-区(1~11 AA),信号肽H-区(12~25 AA)和信号肽C-区(26~33 AA). 根据Phobius预测结果显示34~86氨基酸残基构成非胞质区域(non-cytoplasmic region). 整个GmDIBL1蛋白质序列都比较保守,其中N-端和C-端的保守性更高(图 2b). 系统进化分析结果显示GmDIBL1与棉花、拟南芥的亲缘关系较近,而与水稻、玉米等单子叶植物的B12D的亲缘关系较远(图 3). 这些结果说明GmDIBL1是植物中G-box结合蛋白基因的同源基因,高度的序列保守性暗示其具有相似的生物功能. -
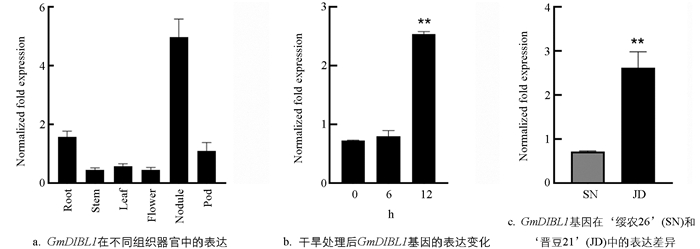
基因的表达特征是分析基因功能的重要数据. 为了明确GmDIBL1基因的表达特征,利用RT-qPCR检测了该基因在大豆不同组织器官中的表达水平、在干旱处理后的表达变化以及耐旱大豆品种(‘晋豆21’)[19]和不耐旱大豆品种(‘绥农26’)[20]中的表达差异. 结果如图 4所示,GmDIBL1基因在大豆的各个组织器官都有表达,在根瘤中的表达水平远远高于其他组织和器官,其次在根和荚中的表达水平较高,在茎、叶、花中的表达水平较低(图 4a). PEG6000处理6 h后,该基因在根中的表达水平有所上升,在处理12 h后,该基因的表达水平显著提高(图 4b). ‘晋豆21’是一个耐旱性较强的大豆品种,而且是一个对干旱比较敏感的品种,GmDIBL1基因在‘晋豆21’根中的表达水平显著高于‘绥农26’根中的表达水平(图 4c). 这些结果说明GmDIBL1基因是一个根、根瘤和荚优势表达基因,受干旱诱导表达,可能在抗/耐干旱胁迫中具有重要作用.
-
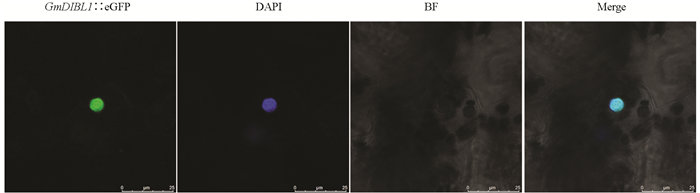
为了分析GmDIBL1的亚细胞定位特征,构建了超量表达GmDIBL1∷eGFP融合基因的植物表达载体并进行了烟草叶片瞬时表达实验,利用激光共聚焦显微镜观测了GFP信号的定位情况. 结果显示,GmDIBL1∷GFP信号与DAPI染色信号重叠,DAPI为细胞核染料,说明GmDIBL1具有细胞核定位(图 5).
-
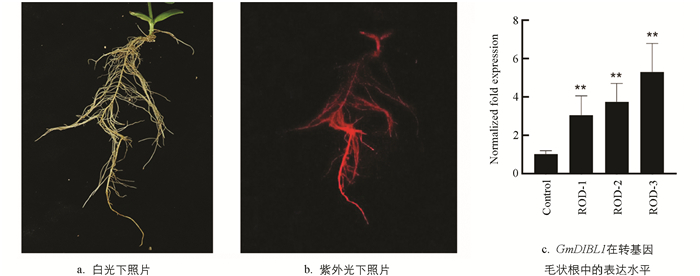
为了获得超量表达GmDIBL1的转基因植物材料,通过构建超量表达GmDIBL1基因的植物表达载体并转入发根农杆菌K599,进而进行大豆毛状根遗传转化. 由于植物表达载体的T-DNA区段中包含CaMV 35S启动子控制GmDIBL1∷eGFP融合基因表达单元,因此首先通过荧光信号观测确定转基因毛状根,并保留1条转基因毛状根以进行后续的研究,结果显示,转基因毛状根在紫外光下能够看到明显的荧光信号(图 6a、6b). 进一步检测GmDIBL1基因在转基因毛状根中的表达变化,结果显示,转基因毛状根中GmDIBL1基因的表达水平都显著高于对照(图 6c). 这些结果说明本研究获得超量表达GmDIBL1基因的转基因毛状根.
-
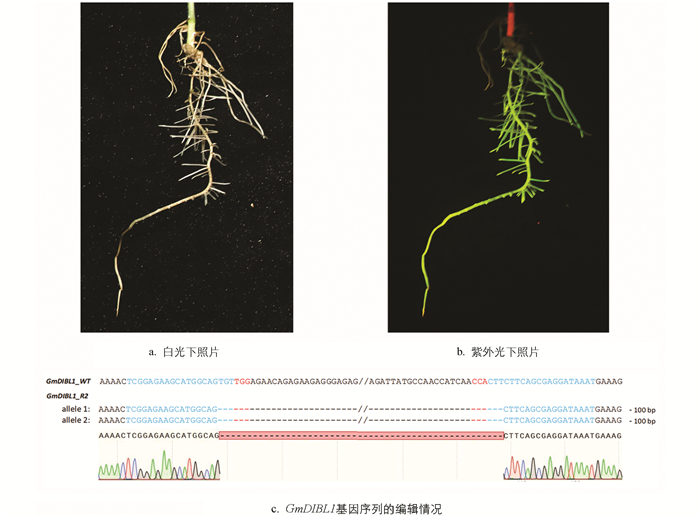
为了获得敲除GmDIBL1的植物材料,通过构建GmDIBL1基因编辑的植物表达载体并转入发根农杆菌K599,进而进行大豆毛状根遗传转化. 由于该载体T-DNA区段内包含1个CaMV35S启动子控制eGFP的表达单元,因此首先观测了毛状根的荧光信号. 结果显示,部分毛状根有明显的荧光信号,说明获得T-DNA区段整合进大豆基因组并正常表达的转基因毛状根(图 7a、7b). 每个复合体植株保留1条转基因毛状根继续培养后进行基因编辑结果分析(图 7c).
-
为了明确GmDIBL1基因在抗/耐干旱胁迫中的功能,利用超量表达GmDIBL1基因和敲除该基因的大豆复合体材料进行干旱耐受性实验. 结果显示,15%的PEG6000处理24 h后,超量表达GmDIBL1基因的毛状根复合体没有萎蔫,而空载对照复合体(CK1和CK2)和基因敲除毛状根复合体出现明显的萎蔫(图 8). 这一结果说明GmDIBL1基因在抗/耐干旱胁迫中具有重要功能,提高该基因表达能够提高大豆耐旱性.
2.1. GmDIBL1基因受干旱处理诱导表达
2.2. GmDIBL1的克隆和序列分析
2.3. GmDIBL1的表达分析
2.4. GmDIBL1亚细胞定位分析
2.5. 超量表达GmDIBL1转基因毛状根的鉴定
2.6. GmDIBL1敲除毛状根的鉴定
2.7. 复合体的抗旱性鉴定
-
在筛选大麦糊粉层和胚的转录组中首次发现了B12D蛋白[12]. B12D家族蛋白是包含参与膜运输的B12D结构域的跨膜蛋白. 通常情况下,B12D蛋白的长度在86和98 AA之间,包括单个跨膜螺旋. 基序预测和多序列比对显示11种开花植物的B12D蛋白具有较强的保守性[18]. 本研究中获得的受干旱诱导的B12D同源蛋白GmDIBL1长度为86个氨基酸残基,具有1个跨膜螺旋,含有B12D结构域,与拟南芥、棉花、番茄和玉米的B12D序列高度同源. 这说明GmDIBL1是大豆中B12D的同源基因. 已有的研究报道显示B12D能够响应多种环境胁迫,例如地黄(Rehmannia glutinosa)B12Dg受铬、GA和盐胁迫诱导表达[11],小麦B12Dg在干旱胁迫下表达下调[15],甜菜(Beta vulgaris)B12Dg能够响应盐胁迫[16],5种水稻基因型不同的水稻中B12Dg都能够响应淹水而上调表达[17]. 生物信息学的分析表明,多数B12D启动子中具有对光、脱落酸、茉莉酸甲酯、细胞分裂素、干旱和热的各种反应元件,少数B12D基因的启动子中存在冷应激、细胞周期、昼夜节律、生长素、水杨酸和赤霉酸的特定元素[6]. GmDIBL1基因能够响应干旱胁迫而显著上调表达,与其他物种同源基因的诱导表达特征相似. 这在表达响应水平上进一步说明GmDIBL1是B12D的同源基因.
-
现有报道中,尽管对B12D基因的诱导表达特征进行了较多的研究,但对B12D功能的研究较少. 本研究中,由于GmDIBL1基因在根瘤和根中优势表达,利用大豆毛状根体系检测了该基因在抗/耐旱中的功能. 在根中超量表达GmDIBL1基因能够显著提高大豆毛状根复合体的抗旱性,但在敲除该基因的复合体中,抗旱性的降低不明显,这一方面可能是PEG6000的使用浓度过高,没有体现出对照和敲除复合体在抗性方面的差异,另一方面可能是因为大豆是多倍体,存在基因功能冗余的现象,单独敲除一个基因难以在表型上出现差异. 本研究正在创制超量表达GmDIBL1以及敲除多个DIBL1基因的大豆材料,以便深入探索该基因的功能和提供抗旱新种质.
3.1. GmDIBL1是植物特异的B12D基因的同源基因
3.2. GmDIBL1在大豆抗/耐干旱胁迫中具有重要作用
-
GmDIBL1(Drought Induced B12D Like 1)是一个受干旱诱导的G-box结合蛋白基因,该基因编码86个氨基酸残基,推测分子量为10.1 kD,等电点为9.647. 推导蛋白质序列与其他物种的G-box结合蛋白之间具有较高的保守性,包含B12D结构域和跨膜螺旋,N-端具有信号肽,C-端构成非胞质区域,并定位于细胞核. GmDIBL1基因在根瘤和根中优势表达,在耐旱品种‘晋豆21’根中的表达水平显著高于干旱敏感品种‘绥农26’. 超量表达GmDIBL1基因的毛状根复合体对干旱胁迫的耐受性增强,证明GmDIBL1在大豆抗/耐干旱逆境中具有重要作用. 该研究为深入解析GmDIBL1介导大豆干旱抗性的调控机制奠定了基础.