


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
可持续农业发展及化肥的减量增效管理是备受关注的全球性议题,与此相应,“菜篮子”工程作为保障居民食物供给、提升膳食质量的重要民生工程,在我国农业转型中具有关键意义。蔬菜含有多种营养物质,是人们维持生命和健康的必需消费品,其品质和产量对我们日常生活非常重要。云南气候独特,区位优势明显,蔬菜以绿色、生态、反季节冬早蔬菜为特点。瓢菜和黄白菜作为昆明市主要栽培的蔬菜,是中国消费者普遍食用和流行的蔬菜[1-2],其生长周期较短,利于进行盆栽试验。研究其不同施肥条件下的光合及光谱作用,对于了解其生长生产特性、实现绿色生产具有重要意义。
光合作用是植物在光照条件下将无机物合成为有机物并且贮存能量的过程,是自然界中最大的碳吸收通量,在减缓气候变化方面发挥着重要作用[3],其强弱对于植物生长及其抗逆性都具有十分重要的影响[4]。作物产量的形成主要取决于光合系统的规模和效率,即光合性能,因此光合作用也是预测产量和品质的重要指标。大量研究表明蔬菜产量形成与其叶片光合性能、光抑制特性及抗衰老特性等关系密切。作物的生长发育受到土壤养分含量和光合特性的影响,任何一种必需养分的缺乏都会降低产量,而土壤养分含量和光合特性又受到施肥方式的调控[5-6]。因此,合理的施肥量及配置比例成为改善作物光合性能,提高光合速率,进而提高产量的有效措施[7-8]。
由于光合作用对环境因子的响应高度敏感[9],不同生理因素、环境条件导致植物光合特性存在差异[10],而借助反射光谱信息可以及时地监测这种生理生化特性的变化[11]。研究表明:植物光谱特征与植物光合和生理有着密切的关系,植物光谱反射信息可以反映出植物的营养状况和植物所处环境的变化[12-13],可用于估测植物理化参数含量,推断植物光合能力,监测植物生长状况等[14-15]。因此,作物光合光谱特性的研究也成为了作物生理生态领域研究的热点。
关于光合特性的研究,不同学者从不同角度取得了丰硕的成果,如不同施肥模式[16-17]、不同种植模式[18]、不同温度措施[19]、不同灌溉模式[20]、不同胁迫[21]以及不同环境条件下作物的光合特性等诸多研究[3, 22]。然而,从前人的研究中关注瓢菜和黄白菜的研究相对较少,且同时关注其光合和光谱特征及相关性的研究更少。因此,本研究以云南省昆明市主要栽培的瓢菜和黄白菜为试验材料,通过设置不同梯度施肥量进行试验处理,测定对应蔬菜的光合性能和反射光谱信息,深入探讨不同施肥条件下蔬菜生长生理、光合特性、光谱特性的机理,旨在为蔬菜生产节约生产成本,促进农业产业可持续发展,同时,也为大面积监测蔬菜的生长信息和预测产量提供理论依据。
全文HTML
-
试验区位于云南师范大学,地处昆明市呈贡区,滇池东岸,这里属于亚热带季风气候,干湿季分明,气候温暖,四季如春,昼夜温差较大,一年四季都可以看到不同的蔬菜种植。研究选取云南省昆明市主要栽培的蔬菜(瓢菜和黄白菜)为试验材料。瓢菜品种为“金品绿霸青梗菜”,黄白菜为“申荣火箭”一代杂交(F1)品种。选取形态完整、大小一致的种子播种,苗期充分灌溉,植株长至4~6片真叶展开、高度为6~8 cm时,选取其中生长性状一致的幼苗移栽至直径20 cm、高15 cm的塑料圆盆中,每盆3株。定植后施用肥料增加土壤养分,确保施肥水平和时间一致。在1~2周的缓苗期后,确保植株正常生长,开始控制施肥量。根据昆明市菜农日常施肥量,试验设计7种施肥模式:C1(对照处理,不施肥)、C2处理(有机肥0 g/m2,复合肥40 g/m2)、C3处理(有机肥0 g/m2,复合肥50 g/m2)、C4处理(有机肥0 g/m2,复合肥60 g/m2)、C5处理(有机肥22 g/m2,复合肥40 g/m2)、C6处理(有机肥22 g/m2,复合肥50 g/m2)、C7处理(有机肥22 g/m2,复合肥60 g/m2),每种模式设置3组重复。分别在蔬菜幼苗期(冬季1月12日)、生长期(冬季1月25日)、成熟期(冬季2月11日)共进行3次数据测量。
-
在晴朗少云、平静无风的天气条件下,于每日09:00-12:00期间进行测定。为尽量减小天气、光照和测定时间差异对光合作用的影响,所有测定均选择在气象条件基本一致的情况下进行。仪器采用GFS-3000高级光合作用-荧光测量系统(德国WALZ公司生产),测定前调试并对设备进行充分预热,通过设备控制测定条件,设定测前温度为(26±1) ℃,CO2摩尔分数为400 ppm,空气流速为750 μmol/s,光强为1 000 μmol/(m2·s),测定时,等待各个数据稳定后保存数据。为了使不同时期测定结果具有可比性,研究控制测定条件和参数尽量一致,测定两种蔬菜幼苗期、生长期和成熟期的叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等参数。不同施肥梯度中,选择不同种类、不同时期且具有代表性的健康成长的植株5株,分别从每株选取3个叶片(保证选择叶位朝向均一致,叶片完全展开、健康、受光一致),每片叶片测定3组数据。处理数据并分析不同施肥条件对蔬菜光合性能参数的影响。
-
与光合参数测定同期进行,利用美国SVC地物光谱仪(HR-1024i),测定蔬菜反射光谱信息。测定均在晴朗少云天气进行,时间为11:00-15:00,同样选择每株蔬菜3个叶片,每片记录10次数据。统计分析不同施肥处理下蔬菜光谱特征变化。
-
用LED红蓝光源设定在梯度光照强度下,测定蔬菜净光合作用速率,拟合光合-光响应曲线。同样设定测前温度为(26±1) ℃,CO2摩尔分数为400 ppm,空气流速为750 μmol/s,叶室中光强由弱到强依次设置为0、30、50、100、200、300、400、600、800、1 000、1 200、1 400、1 600、1 800和2 000 μmol/(m2·s)。在测定时,使蔬菜适应每一梯度光强3~5 min。测定计算所得参数包括:暗呼吸速率(Rday)、光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Amax)及表观量子效率(AQE)等。
-
应用Excel 2013、Origin 2022、SPSS 22、和SVC HR-1024i配套软件,计算试验数据并作图,计算归一化植被指数(Normalized Difference Vegetation Index,NDVI)[23],氮素反射率指数(Nitrogen Reflectance index,NRI)[24],比值植被指数(Ratio Vegetation Index,RVI)[25],差值植被指数(Difference Vegetation Index,DVI)[26]等相关光谱特征参数(表 1),以深入探讨不同施肥条件下蔬菜叶绿素光合、光谱特性与施肥条件的相关性。
1.1. 试验设计
1.2. 蔬菜光合参数测定
1.3. 蔬菜反射光谱信息测定
1.4. 蔬菜光合-光响应曲线测定
1.5. 数据处理
-
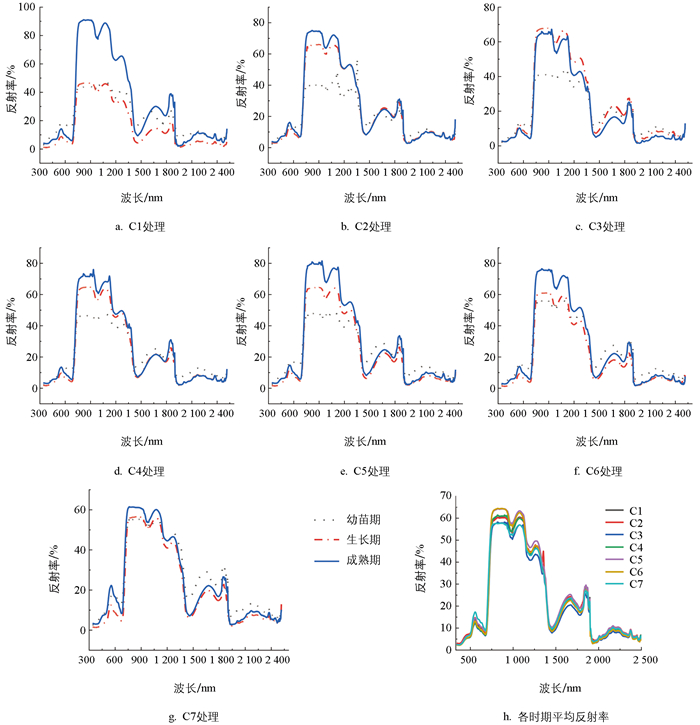
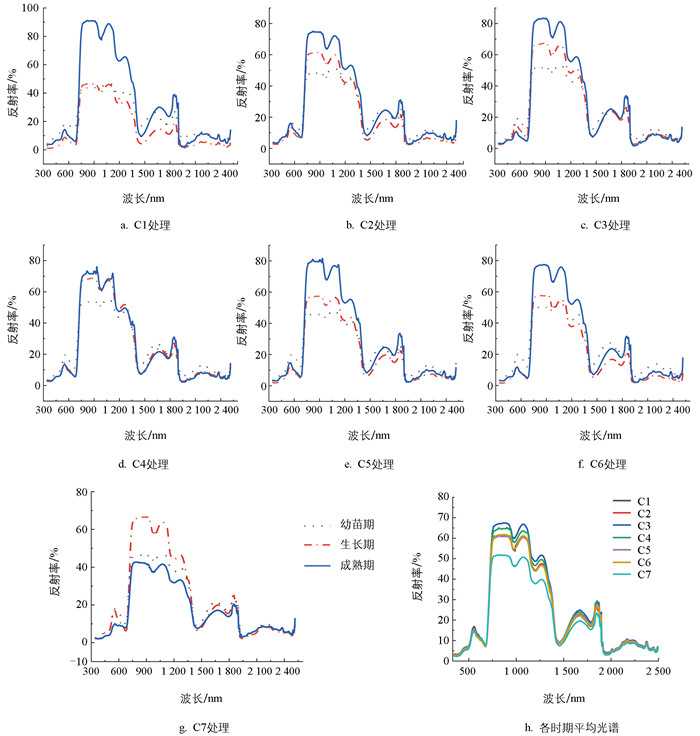
由于蔬菜在不同生长时期的生长量和生长发育速度不同,对营养条件的要求也不同,因此不同生育期不同施肥条件下蔬菜光谱反射率具有一定差异,但总体上瓢菜和黄白菜均与绿色植物叶片光谱特征相似。在可见光区域(350~700 nm)形成两个叶绿素吸收谷和绿色反射峰;在近红外区域(800~1 300 nm)形成高反射率红肩;在红光波段与近红外区域之间(700~800 nm)形成“红边”;在短波红外区域(1 300~2 500 nm),主要为受水分控制形成的吸收谷和反射峰。
具体来看,根据各时期的平均反射率可知,不同施肥处理下瓢菜光谱反射率差异主要体现在反射率较强的绿峰和近红外区域的反射率红肩,以及短波红外区域的水分反射峰处。在绿峰区域C7处理的反射率最高,在近红外区域(800~1 300 nm)反射率较低的主要为C3处理和C7处理,而反射率较高的主要为C5和C6处理,在短波红外区域(1 300~2 500 nm)C3处理的反射率依然较低,C5处理的反射率较高。从不同生育期来看,其差异主要体现在800~1 300 nm的近红外区域,除C3处理,7种不同处理模式下成熟期的反射率均较幼苗期和生长期高。其中C1未施肥对照处理模式下幼苗期和生长期反射率相差不大,随着施肥量的增加,幼苗期反射率变化不大,C6处理和C7处理的反射率有所提高;而生长期从C2处理开始反射率明显提高,从40%左右提高到60%左右;成熟期的反射率反而随着施肥量的增加有所降低,C7处理模式下不同生育期的光谱反射率相差较小(图 1)。
黄白菜在不同施肥处理不同生育期光谱特征与瓢菜具有相似性又有差异性,其中C1未施肥处理下两种蔬菜光谱反射率差异不大。由图 2可知,在不同施肥处理下黄白菜光谱反射率差异主要体现在800~1 300 nm的近红外区域和1 550~1 890 nm的短波红外区域,主要表现为C3处理反射率较高,C7处理反射率较低。不同施肥处理各生育期反射率均存在差异,C1到C6处理下反射率以成熟期较高,而C7处理变为生长期反射率较高,C4处理下生长期和成熟期光谱反射率较为接近。
-
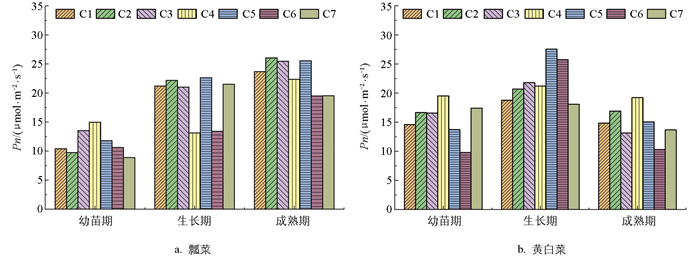
光合速率是度量作物源强(源器官同化物形成和输出光合产物的总能力)最直观的一个指标。通过测定瓢菜和黄白菜净光合速率值(Pn),并绘制不同施肥处理下净光合速率变化图(图 3)。可以看出,瓢菜净光合速率随生育期的推进而明显升高,除C4施肥模式下生长期Pn值较幼苗期和成熟期低,其他施肥模式下瓢菜不同生育期Pn值从大到小依次为成熟期、生长期、幼苗期;随施肥水平的提高瓢菜净光合速率在幼苗期和成熟期呈现先增加后降低的趋势。Pn值最高为成熟期的C2处理,达到26 μmol/(m2·s);最低为幼苗期C7的处理,Pn值为8.8 μmol/(m2·s)。
对比瓢菜,黄白菜Pn值随生育期的推进呈先增高后降低的趋势,生长期的净光合速率Pn值大于幼苗期和成熟期,其中C5、C6处理下生长期Pn值较幼苗期增长较多。随施肥水平的提高,黄白菜Pn值在成熟期C5、C6、C7处理下表现出明显的抑制作用,尤其C6处理下黄白菜Pn值最低。说明适当施肥对黄白菜生长发育前期Pn值有明显的提高作用,较高施肥量则对成熟期黄白菜Pn值表现出一定的抑制作用。
-
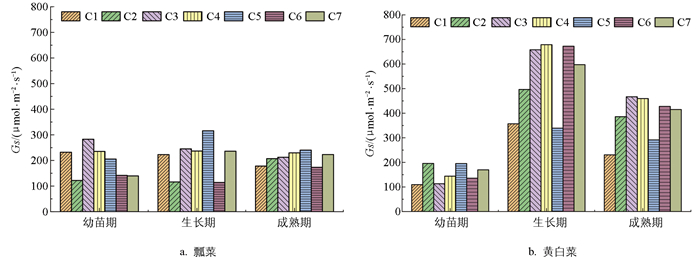
图 4为不同施肥处理下瓢菜和黄白菜气孔导度(Gs)的对比图,从中可以看出,随着生育期的推进和施肥水平的提高,瓢菜Gs值呈不同的变化趋势。生长期C5处理的Gs值最高,为315.9 μmol/(m2·s);生长期C6处理的Gs值最低,为113.9 μmol/(m2·s)。C3、C4、C5处理Gs值整体上处于较高水平,C2、C6处理Gs值明显较低。
不同施肥处理下,黄白菜生长期Gs值大于幼苗期和成熟期。生长期C4处理的Gs值最高,为678.7 μmol/(m2·s);幼苗期C1处理的Gs值最低,为109.5 μmol/(m2·s)。在生长期和成熟期,黄白菜Gs值随施肥水平的提高呈先增加后降低的趋势,C5处理下Gs值明显降低,与瓢菜相比,施用肥料能显著提高黄白菜各生育时期的Gs值。
-
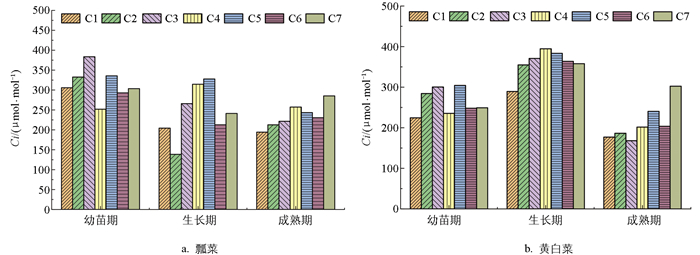
图 5为不同施肥处理下瓢菜胞间CO2浓度(Ci)的对比图,从中可以看出,随生育期的推进,瓢菜Ci值逐渐下降,除了C4处理,其他施肥处理下均是幼苗期的Ci值大于生长期和成熟期。幼苗期C3处理的Ci值最高,为383.6 μmol/mol;生长期C2处理的Ci值最低,为138.5 μmol/mol。
相比瓢菜,黄白菜Ci值在不同施肥处理下生长期大于幼苗期和成熟期,即随生育期的推进,黄白菜胞间CO2浓度先增加后降低。生长期C4处理的Ci值最高,为394.6 μmol/mol;成熟期C3处理的Ci值最低,为168.1 μmol/mol。
-
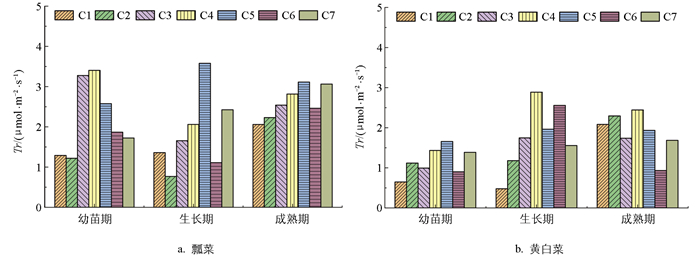
图 6为冬季不同施肥处理下瓢菜蒸腾速率(Tr)的对比图,从中可以看出,瓢菜和黄白菜Tr值随生育期的推进而呈现出不同的变化趋势。瓢菜生长期C5处理的Tr值最高,为3.6 μmol/(m2·s);生长期C2处理的Tr值最低,为0.8 μmol/(m2·s)。随施肥水平的提高,瓢菜幼苗期Tr值先升高后降低,生长期C2、C6处理下Tr值较低,C5处理下Tr值最高,成熟期时C5、C7处理下Tr值较高。
黄白菜生长期C4处理的Tr值最高,为2.9 μmol/(m2·s);生长期C1处理的Tr值最低,为0.4 μmol/(m2·s)。生长期C4、C6处理的Tr值较高,成熟期C2、C4处理的Tr值较高。与瓢菜相比,黄白菜总体Tr值处于较低水平。生长期时适量施肥能明显提高黄白菜Tr值,成熟期足量施肥反而有降低Tr值的作用。
-
非直角双曲线模型是目前拟合光合-光响应曲线较普遍使用的模型[27],由于模型本身的特点,光合光响应曲线是一条趋近于极值的渐近线[28]。非直角双曲线模型公式如下。
式中:Pn为净光合速率;AQE为表观量子效率;PAR为有效光合辐射;Amax为最大净光合速率;Rday为暗呼吸速率;K为光响应曲线曲角。在设置AQE、Amax、Rday、K初始值后,运行SPSS 22软件进行迭代运算,即可得到模型模拟值[29-30]。
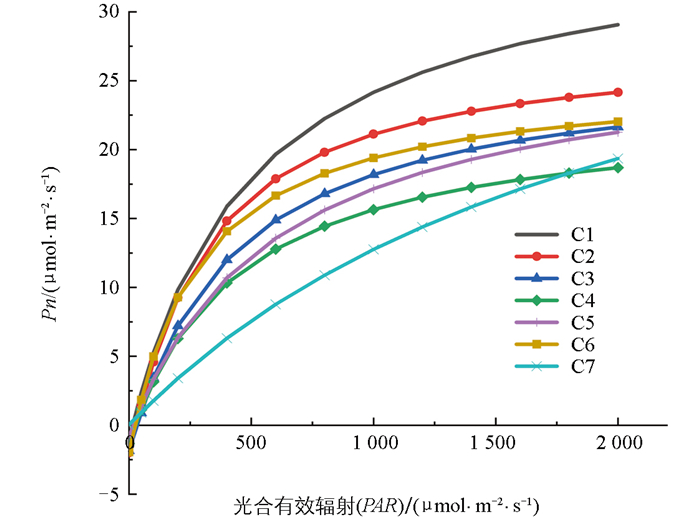
从图 7中可知,非直角双曲线模型拟合得到的净光合速率(Pn)变化趋势整体上是随光合有效辐射(PAR)增强而增大的,增幅随着PAR的增强而减缓,在达到光饱和点后Pn值迅速下降,平均R2为0.944,拟合效果较好。
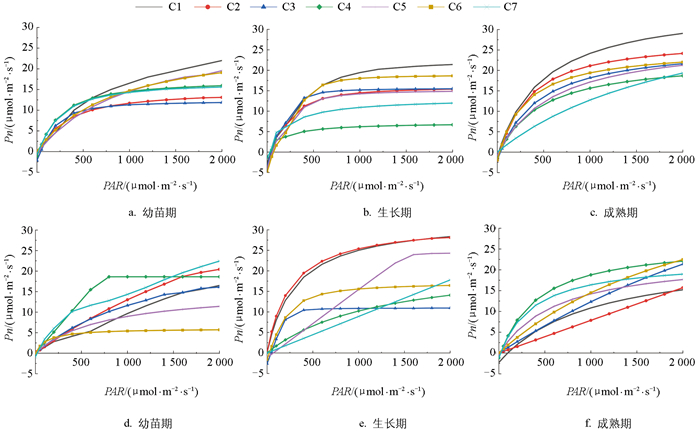
从图 8可以看出,不同施肥处理下瓢菜和黄白菜Pn对光强具有不同响应趋势。整体来看,瓢菜的Pn值变化趋势是先随光合有效辐射(PAR)增强而迅速增大,增幅随着PAR的增强而减缓,在达到光饱和点后Pn值迅速下降。随着生育期的推进,光响应曲线的峰值逐渐增加。在试验设定的光合有效辐射范围内,7种处理均未出现光抑制现象。黄白菜的光合-光响应曲线比瓢菜变化大,幼苗期C4处理、生长期C1、C2处理、成熟期C4处理的Pn先随PAR增强而迅速增大,在达到光饱和点后Pn变化不大,且其最大净光合速率在所有处理中最大;幼苗期C6处理、生长期C6处理、成熟期C5处理的Pn在PAR较低光强下就出现光饱和。利用非直角双曲线对两种蔬菜进行拟合的整体效果较好,在有效光合辐射(PAR)处于较低水平,即PAR<200 μmol/(m2·s)时,瓢菜和黄白菜Pn增长较快,说明在低PAR下蔬菜利用光能的效率较高。从光合-光响应曲线可知,合理配置施肥模式可以获得更高效益。
-
不同施肥条件下瓢菜、黄白菜光合参数与光谱植被指数的相关系数见表 2。瓢菜最大正相关系数为0.880,说明C7处理的NRI与Ci呈显著正相关;最小负相关系数为-0.961,说明C5处理的NDVI与Ci呈显著负相关。黄白菜最大正相关系数为0.880,说明C3处理的NRI与Ci呈显著正相关;最小负相关系数为-0.930,说明C5处理的NDVI与Gs呈显著负相关。此外,瓢菜比值植被指数RVI在C2处理下与光合参数的相关性均大于0.5,C5处理下归一化植被指数NDVI与光合参数的相关性均大于0.6,C4处理下瓢菜氮素反射指数NRI与光合参数的相关性均大于0.5,且C4处理下差值植被指数DVI与两种蔬菜光合参数的相关性均大于0.8,说明C4处理下有利于提高两种蔬菜光合性能,且瓢菜不同施肥处理下光谱指数与光合参数的相关性整体上较黄白菜更好。不同施肥条件蔬菜光合生理状况存在差异,蔬菜植被指数与光合参数存在显著相关性,因此,可利用蔬菜对不同施肥条件的光谱特征和光合性能的响应来监测蔬菜生长状况。
2.1. 蔬菜光谱反射率强度变化
2.2. 蔬菜光合作用强度变化
2.2.1. 蔬菜净光合速率(Pn)
2.2.2. 蔬菜气孔导度(Gs)
2.2.3. 蔬菜胞间CO2浓度(Ci)
2.2.4. 蔬菜蒸腾速率(Tr)
2.3. 蔬菜光合-光响应特性变化
2.4. 蔬菜光合性能与光谱植被指数相关性
-
根据结果分析可知,不同施肥条件和生育期对两种蔬菜的光谱反射率和光合特征都有影响。在C1未施肥处理下两种蔬菜光谱反射率差异不大,总体上表现为成熟期的光谱反射率最大;而不同施肥模式下两种蔬菜的光谱反射率发生了不同的变化。这说明不同施肥模式使土壤中营养元素含量发生变化,进而影响蔬菜的生长发育。由于两种不同蔬菜叶片表面特性和内部结构以及营养成分浓度和分布都有差异,因此在不同施肥模式下光谱变化特征也具有差异[31]。在不同生育期的光谱差异主要体现为:在生长期,瓢菜在C1施肥模式下各波段反射率均较低,在可见光和短波红外区域C2施肥模式反射率较高,在近红外区域C3施肥模式反射率较高;而黄白菜生长期光谱反射率较低的为C6施肥模式,在绿峰和近红外区域C1施肥模式反射率较高,在短波红外区域C3施肥模式光谱反射率较高。在成熟期,瓢菜在可见光区域C7处理模式光谱反射率较高,C3处理模式反射率较低,在近红外和短波红外区域C1施肥模式光谱反射率较高,C3和C7处理模式光谱反射率较低;黄白菜成熟期C7处理模式光谱反射率明显变低,在可见光区域C2施肥模式光谱反射率较高,在近红外区域C3和C5施肥模式光谱反射率较高。说明适量的肥料能加快作物生长发育,但当施肥水平超过一定值时,反而抑制作物生长,光谱反射率降低[7]。
不同施肥条件下两种蔬菜光合性能差异主要表现为:净光合速率(Pn)值除C4和C6施肥处理模式以外,均是瓢菜大于黄白菜,瓢菜净光合速率随生育期的推进而明显升高,黄白菜净光合速率随生育期的推进呈先增高后降低的趋势。Li等[32]类似的研究表明,随着硅酸钾肥用量的增加,小麦旗叶叶绿素含量、净光合速率呈先增加后减少的趋势,说明适当施肥对黄白菜生长发育前期净光合速率有明显的提高作用,较高施肥量则对成熟期黄白菜表现出一定的抑制作用。蒸腾速率(Tr)值除C2施肥处理模式,其他均是瓢菜大于黄白菜,生长期时适量施肥能明显提高黄白菜蒸腾速率,成熟期足量施肥反而有所降低蒸腾速率。胞间CO2浓度(Ci)值两种蔬菜相差不大,说明施肥对提升两种蔬菜胞间CO2浓度的作用不明显。7种施肥模式气孔导度(Gs)值均表现为黄白菜大于瓢菜,不同施肥处理下瓢菜气孔导度提升不显著,而黄白菜各生育时期的气孔导度得到显著提高。冯茜等[17]的研究表明气孔导度、胞间CO2浓度和蒸腾速率都与光合速率显著相关。Fu等[33]的研究表明光合速率的准确表征对于重新设计光合作用途径以提高作物产量具有关键价值。因此,可以通过施肥调整这些光合参数来提高叶片光合速率,为提高蔬菜产量提供参考。
此外,光合-光响应曲线可以帮助我们理解光合机制,评估植物如何对光照条件做出反应,或评估植物内部生理可塑性的程度[34]。本研究表明在有效光合辐射(PAR)处于较低水平,即PAR<200 μmol/(m2·s)时,瓢菜和黄白菜净光合速率增长较快,说明在低PAR下蔬菜利用光能的效率较高。Babla等[35]的研究也表明与受弱光和中光照的植物相比,植物在长期强光下的光合作用潜力会受到影响,而从光合-光响应曲线可知,合理施用肥料则可以提高蔬菜对强光的利用,获得更高效益。
-
不同施肥条件下瓢菜和黄白菜光谱特征参数与光合性能间相关性也具有差异。瓢菜C7处理的NRI与Ci呈显著正相关,C5处理的NDVI与Ci显著负相关;黄白菜C3处理的NRI与Ci显著正相关,C5处理的NDVI与Gs显著负相关。这说明两种蔬菜的同一光谱指数与同一光合参数和不同光合参数的相关性在不同施肥模式下均具有差异,这可能是蔬菜生理结构特征的差异造成的。C4处理下差值植被指数DVI与两种蔬菜光合参数的相关性均大于0.8,说明C4处理下有利于提高两种蔬菜光合性能,且瓢菜不同施肥处理下光谱指数与光合参数的相关性整体上较黄白菜更好。Peñuelas等[31]的研究表明植物光合作用能力是反映植物生长状况和适应逆境胁迫的重要指标,而植物在生长过程中的微小变化可以通过植物光谱特征表现出来,其研究结果表明归一化植被指数(NDVI)、红边参数(λre)、光化学反射指数(PRI)等与其他光合效率指标之间存在普遍的相关性。也有研究表明植物叶片不同生长阶段的光响应程度具有差异,如Sánchez-de-Miguel[36]等的研究表明成熟叶片光合作用的光响应高于年轻叶片和老叶片的光响应。因此,可应用蔬菜对不同施肥条件的光谱特征和光合性能的响应来监测蔬菜生长状况,从而促进蔬菜的科学管理和产量的提高。
-
研究利用GFS-3000便携式光合-荧光测量系统和SVC便携式地物光谱仪对瓢菜和黄白菜的光合及光谱特性进行测量,主要结论如下:
1) 不同施肥条件和生育期对两种蔬菜的光谱反射率有很大影响,瓢菜反射率除C3处理外,其他处理模式下成熟期的反射率均较幼苗期和生长期高;黄白菜C1到C6处理下反射率以成熟期较高,而C7处理生长期反射率较高。
2) 7种施肥模式下,瓢菜和黄白菜的光合性能表现出明显差异。瓢菜叶片净光合速率(Pn)随生育期的推进而明显升高,黄白菜Pn随生育期的推进呈先增高后降低的趋势;瓢菜叶片蒸腾速率(Tr)最高为生长期C5处理,黄白菜最高为生长期C4处理;两种蔬菜叶片胞间CO2浓度(Ci)波动不大;气孔导度(Gs)在不同施肥处理下瓢菜提升不显著,黄白菜Gs则显著提高;光合-光响应曲线表明在有效光合辐射(PAR)处于较低水平,即PAR<200 μmol/(m2·s)时,瓢菜和黄白菜Pn增长较快。
3) 不同施肥条件下蔬菜光合生理状况存在差异,蔬菜光谱特征参数与光合性能间相关性也不同。瓢菜C7处理的氮素反射率指数NRI与Ci呈显著正相关,C5处理的归一化植被指数NDVI与Ci呈显著负相关;黄白菜C3处理的NRI与Ci呈显著正相关,C5处理的NDVI与Gs呈显著负相关。
研究选取的蔬菜为昆明地区常见设施蔬菜中的两种,受试验条件限制,为不覆盖塑料薄膜种植,但昆明地区蔬菜产业多为大棚设施种植,二者光谱信息存在差异,以后的试验应尽量还原大棚种植环境,进一步贴合蔬菜实际生产状况。此外,本试验数据仅为一季数据,所以其结果仍需在不同的田间环境下加以验证和完善。