


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
随着全球气候变暖加剧和肉鸡养殖向高密度、集约化饲养模式的转变,热应激(Heat stress)成为影响家禽业可持续发展的主要环境因素之一[1-2]。由于肉鸡具有羽毛丰厚、皮肤缺乏汗腺、代谢旺盛、体温高等特点,其机体状态和生产性能极易受到环境温度的影响[3]。研究表明,肉鸡在环境温度超过32 ℃时容易出现热应激反应[4],包括体温升高、生长性能下降、肠道结构和功能受损、肠道微生物群落结构的改变[5]、机体氧化应激及全身炎症反应等,严重影响肉鸡饲养的经济效益。
肠道作为营养物质消化和吸收的主要场所[6],其结构和功能的完整性在家禽生长发育过程中至关重要[7]。健康的肠道依赖于物理、化学和免疫屏障的协同作用,其中,肠绒毛的形态、紧密连接蛋白的表达以及杯状细胞分泌的黏液共同构成了抵御病原体和毒素入侵的第一道防线。有研究表明,肠道是热应激反应的中心器官[8]。当家禽处于持续高温时,机体出于自我保护会重新分配体内血流量,减少流经肠道的血流量[9],导致肠道严重缺血缺氧、上皮细胞脱落、肠道屏障功能被破坏。肠道黏膜屏障的损伤会增加肠道通透性,使有害微生物和毒素更容易进入血液循环[10],造成全身性的炎症反应,进一步加重组织损伤。
肠道微生物群在动物健康和疾病状况中起着重要作用[11]。肠道菌群参与营养物质的代谢、屏障功能的维持以及免疫调节等多种生理过程,能通过与肠上皮细胞表面结合形成生物屏障,阻止病原菌的定植与侵袭[12]。研究表明,高温环境对肠道造成的损害与肠道微生物有关[13]。热应激能够影响肠道菌群的组成和功能,导致肠道菌群失调,具体表现为有害菌过度增殖、有益菌丰度降低,进一步造成肠上皮细胞损伤、紧密连接蛋白表达下调、黏膜屏障完整性损坏,从而影响家禽的健康和生长性能[14-15]。
本研究以AA肉鸡为研究对象,建立了为期21 d的慢性热应激模型,从生长性能、肠道形态结构、屏障功能、氧化应激、炎症水平以及肠道菌群生态等多个维度,研究慢性热应激致肉鸡肠道损伤的病理生理机制,旨在探究慢性热应激致肉鸡肠道损伤的作用机制,揭示“氧化应激-屏障损伤-炎症反应-菌群失衡”这一恶性循环通路,为研究和开发缓解热应激的营养策略和生物技术手段提供理论依据,对保障家禽健康和提高生产性能具有重要的实践意义。
全文HTML
-
皮质酮(Corticosterone,CORT)、去甲肾上腺素(Norepinephrine,NE)、白细胞介素-1β(Interleukin-1β,IL-1β)、白细胞介素-6(Interleukin-6,IL-6)、肿瘤坏死因子-α(Tumor Necrosis Factor-α,TNF-α)等酶联免疫吸附测定(Enzyme-Linked Immunosorbent Assay,ELISA)检测试剂盒购于武汉云克隆科技股份有限公司;过氧化氢酶(Catalase,CAT)、丙二醛(Malondialdehyde,MDA)、超氧化物歧化酶(Superoxide Dismutase,SOD)活性检测试剂盒、4′,6-二脒基-2-苯基吲哚(4′,6-diamidino-2-phenylindole,DAPI)溶液、中性树胶购于北京索莱宝科技有限公司;谷胱甘肽过氧化物酶(Glutathione Peroxidase,GSH-Px)、总抗氧化能力(Total Antioxidant Capacity,T-AOC)检测试剂盒购于南京建成生物工程研究所;苏木素-伊红(Hematoxylin-Eosin,HE)染色液购于白鲨生物有限公司;过碘酸-希夫(Periodic Acid-Schiff,PAS)染色试剂盒购于索莱宝生物有限公司;末端脱氧核苷酸转移酶介导的dUTP缺口末端标记(Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick End Labeling,TUNEL)免疫荧光试剂盒购于赛维尔生物有限公司;光学显微镜购自日本Olympus公司;石蜡包埋机、切片机、组织摊片机购于德国莱卡公司;艾本德高速低温离心机购于德国Eppendorf公司。
-
本研究严格遵循贵州大学动物试验伦理规范,所有试验操作均经过贵州大学试验动物伦理委员会的批准(批准编号:EAE-GZY-2024-T222)。
试验动物:21日龄雄性AA肉鸡20只,体质量180±20 g,饲养于环境温度22±2 ℃、环境湿度50±5%的试验动物饲养房中。适应7 d后,随机分为对照组(Control组)和热应激组(Stress组),每组10只鸡。热应激组每天进行5 h热应激(9:00-14:00),温度为33±2 ℃,湿度为50±5%,其余时间的温度控制在22±2 ℃。自由采食和饮水,试验持续21 d。试验过程中记录体质量、食物摄入量、体温变化。造模结束后,按照动物伦理审批方案对肉鸡实施安乐死。血液样本肝素钠抗凝后于4 ℃保存,肠组织样本处理后于-80 ℃保存。
-
在试验第1 d和第21 d对肉鸡空腹称重并计算平均日增重(Average Daily Gain,ADG)。试验期间每日记录各组肉鸡的总采食量,计算平均日采食量(Average Daily Feed Intake,ADFI)和料重比(Feed to Gain Ratio,F/G)。
式中:Wf为试验末期平均体质量(g);Wi为试验初期平均体质量(g);t为试验天数(d);It为试验期总采食量(g);n为每组肉鸡数量。
-
试验结束后取肉鸡血液,经4 ℃、3 000 r/min离心10 min收集血浆保存于-80 ℃冰箱中。参照武汉云克隆ELISA检测试剂盒说明书对肉鸡血浆中CORT、NE、IL-6、IL-1β、TNF-α进行检测,在酶标仪450 nm波长处测量各孔吸光度值,按照说明书计算最终浓度。
-
取肉鸡肠道组织用4%多聚甲醛固定液固定,制作成石蜡切片,烘干、脱蜡、并进行浓度梯度酒精复水,经HE染色后封片,于显微镜下镜检组织学变化。使用Image-Pro Plus 6.0软件统计每张图片中肠绒毛高度(Villi,V)和隐窝深度(Crypt,C),再计算肠绒毛高度与隐窝深度比值(V/C)。
-
肉鸡肠道组织经石蜡包埋后制备成组织切片,以1×柠檬酸钠缓冲液(pH 6.0)进行热诱导抗原修复,加热20 min后自然冷却至室温;随后用磷酸盐缓冲盐水(Phosphate-Buffered Saline,PBS)洗涤3次,接着采用封闭液封闭组织切片中的非特异性结合位点;之后分别滴加经PBS体积稀释的一抗(兔源Claudin-1,稀释比例1∶600;兔源Occludin,稀释比例1∶400),于4 ℃环境下孵育12 h。二抗(山羊抗兔IgG-HRP)以PBS按体积稀释,稀释比例为1∶200,于37 ℃孵育1 h;随后采用二氨基联苯胺(3,3-diaminobenzidine,DAB)进行显色,显色后以苏木素染液复染3 min;接着经浓度梯度酒精脱水、二甲苯透明,最后用中性树胶封片,于显微镜下观察并拍照记录。采用Image-Pro Plus 6.0软件计算组织中Claudin-1、Occludin阳性区域平均吸光度值。使用索莱宝PAS染色试剂盒,切片先常规脱蜡复水;经高碘酸溶液氧化,氧化后流水冲洗;滴加雪夫试剂染色,染色后流水冲洗;用试剂盒配套苏木素染液复染,随后经1%盐酸酒精分化、流水返蓝;再经浓度梯度酒精脱水、二甲苯透明,最后用中性树胶封片,于显微镜下拍照记录。
-
将切片进行抗原修复后通透平衡,使用标记工作液进行标记反应1 h,PBS洗涤3次,每次5 min。采用DAPI细胞核染色8 min,抗荧光淬灭封片剂封片,立即在荧光显微镜下观察拍照。采用Image-Pro Plus 6.0软件计算组织中绿色荧光阳性区域平均吸光度值。
-
参照索莱宝、南京建成氧化应激指标活性检测试剂盒说明书,测定肉鸡血浆中CAT、MDA、SOD、GSH-Px及T-AOC水平。
-
将盲肠内容物置于冻存管中做好标记后,液氮冻存,送至上海美吉生物医药科技有限公司在Illumina MiSeq PE300系统上进行高通量测序。
-
使用IBM SPSS 26.0软件对试验数据进行显著性差异分析,通过单因素ANOVA检验组间差异,并进行LSD及邓肯事后检验。组间差异通过字母标记法进行标记,小写字母不同表示组间具有统计学意义(p<0.05),小写字母相同或无字母表示组间不具有统计学意义(p>0.05)。使用GraphPad Prism 10.1.2绘图。
1.1. 主要试剂与仪器
1.2. 试验动物与处理
1.2.1. 生长性能测定
1.2.2. ELISA检测
1.2.3. 肉鸡肠道HE染色
1.2.4. 肉鸡肠道免疫组化和PAS染色
1.2.5. 肉鸡肠道TUNEL染色
1.2.6. 氧化应激指标检测
1.2.7. 肠道菌群高通量测序
1.2.8. 数据统计与分析
-
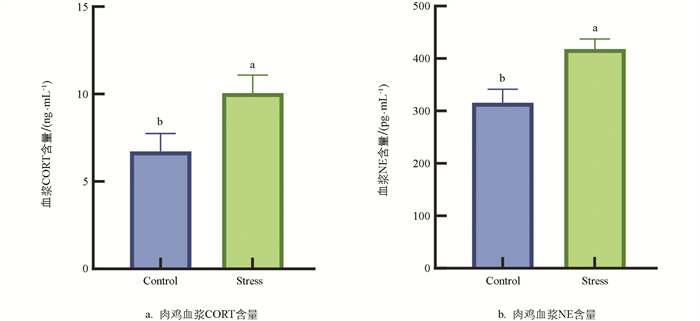
检测两组血浆中应激激素CORT和NE水平,验证机体是否处于应激状态。如图 1所示,与Control组相比,Stress组肉鸡血浆CORT和NE水平显著升高53.35%、32.49%(p<0.05),提示慢性热应激肉鸡模型建立有效。
-
如表 1所示,与Control组相比,Stress组肉鸡的ADG和ADFI显著降低(p<0.05),F/G显著升高(p<0.05),提示热应激显著影响肉鸡生长性能。如表 2所示,通过测量体温,发现在第1 d各组间体温无显著差异(p>0.05);第7、14、21 d测量显示,与Control组相比,Stress组的体温均显著升高(p<0.05)。
-
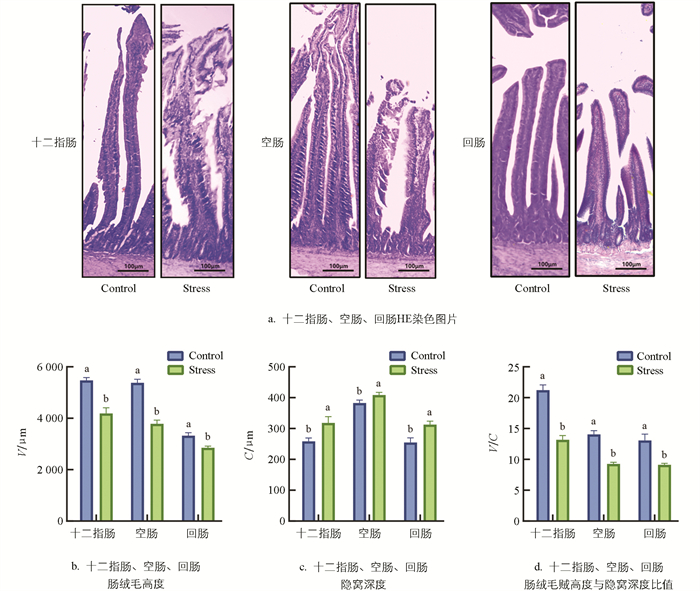
如图 2a所示,小肠组织经HE染色显示热应激导致肉鸡十二指肠、空肠和回肠结构受损,黏膜组织脱落,固有层裸露,肠绒毛断裂;对肠绒毛和隐窝进行测量(图 2b、2c、2d),结果显示,与Control组相比,Stress组肉鸡十二指肠、空肠和回肠肠绒毛V显著降低(p<0.05),C显著增加(p<0.05),V/C显著降低(p<0.05),提示慢性热应激会导致肉鸡肠道物理屏障损伤。
-
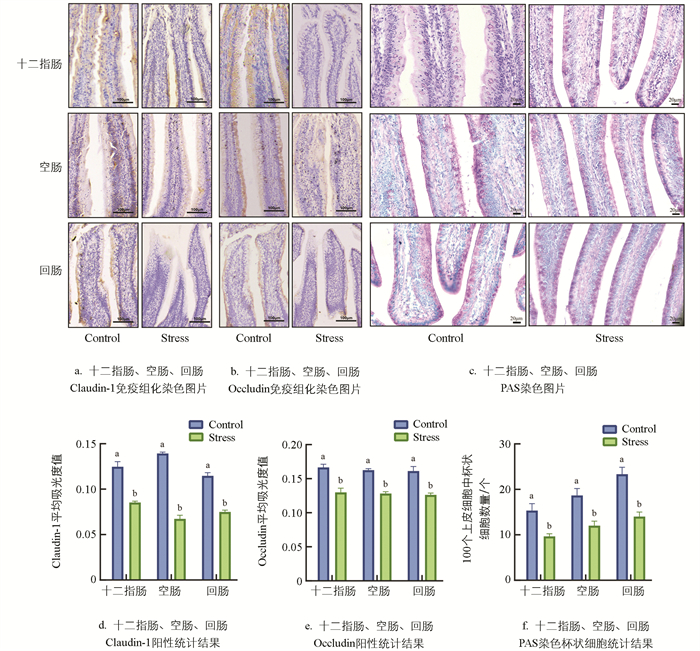
免疫组织化学结果显示,Stress组肉鸡十二指肠、空肠、回肠紧密连接蛋白Claudin-1阳性表达较Control组显著降低31.55%、51.67%、34.59%(p<0.05,图 3a、3d),Occludin阳性表达较Control组显著降低22.00%、21.11%、21.69%(p<0.05,图 3b、3e)。通过PAS染色检测杯状细胞数量,结果显示与Control组相比,Stress组十二指肠、空肠、回肠杯状细胞数量显著减少36.96%、35.71%、39.40%(p<0.05,图 3c,3f)。提示热应激通过减少各肠段中紧密连接蛋白Claudin-1、Occludin的阳性表达量和杯状细胞数量,造成肉鸡肠道屏障损伤。
-
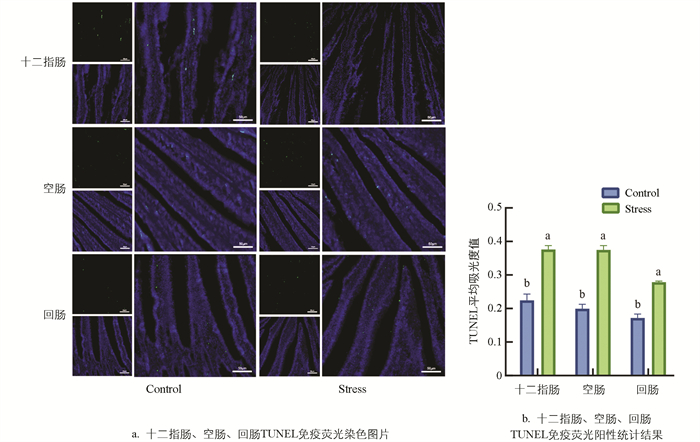
如图 4所示,TUNEL染色后,Stress组肉鸡十二指肠、空肠和回肠TUNEL阳性表达比Control组分别升高53.11%、88.06%、61.91%(p<0.01),提示慢性热应激会导致肉鸡肠道细胞凋亡增加。
-
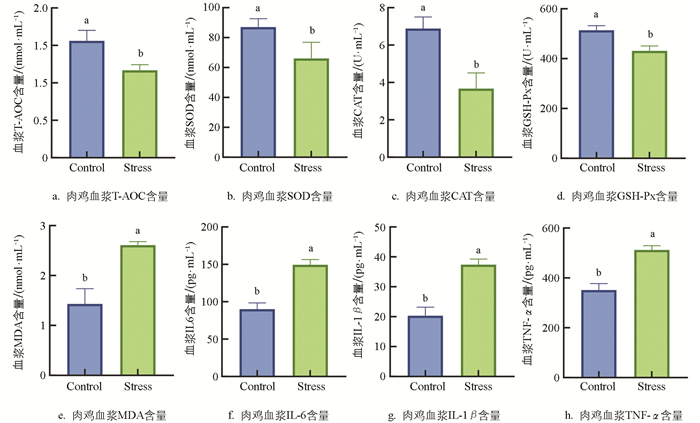
ELISA试验检测显示,与Control组相比,Stress组肉鸡血浆T-AOC、SOD、CAT和GSH-Px含量显著降低17.79%、24.12%、46.67%、16.10%(p<0.05,图 5a、5b、5c、5d),MDA含量显著增加82.16%(p<0.05,图 5e),提示机体处于氧化应激状态,慢性热应激可诱导肉鸡发生氧化应激。
-
与Control组相比,Stress组肉鸡血浆IL-1β、IL-6和TNF-α含量显著上升65.72%、84.25%、45.85%(p<0.05,图 5f、5g、5h),提示慢性热应激会导致机体产生炎症反应。
-
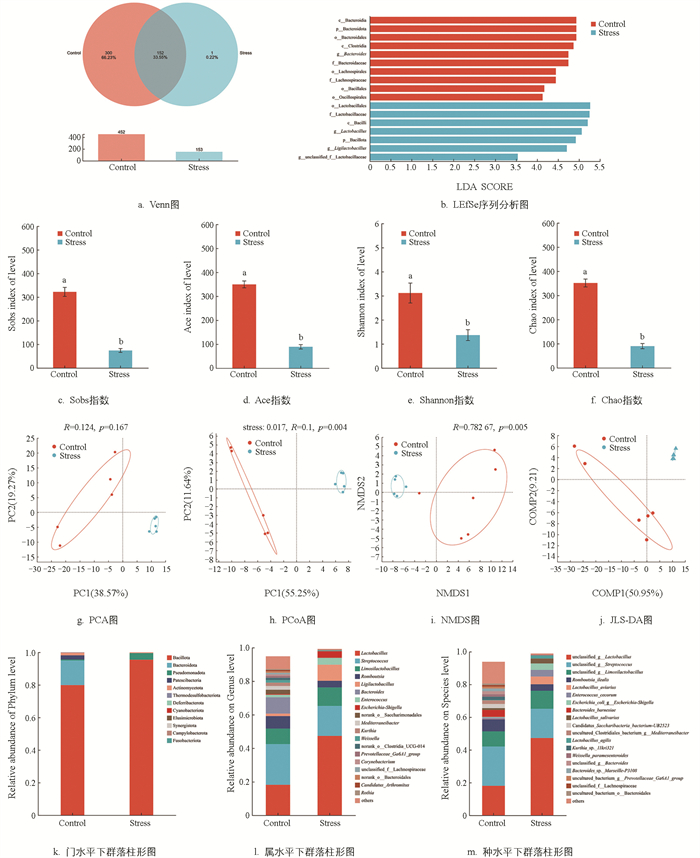
微生物16S rRNA测序分析结果显示,Control组OTU总数显著高于Stress组(p<0.05,图 6a)。菌群的α多样性显示,Stress组的各指数均显著低于Control组(p<0.05,图 6c、6d、6e、6f),提示慢性热应激显著降低肉鸡肠道菌群的丰富度和多样性。菌群的β多样性显示,Control组和Stress组组间主成分完全分离(图 6g、6h、6i、6j),提示慢性热应激显著改变肉鸡肠道菌群的群落结构;物种相对丰度显示,门水平下,与Control组相比,Stress组肉鸡盲肠中厚壁菌门(Bacillota)和变形菌门(Pseudomonadota)相对丰度增加,拟杆菌门(Bacteroidota)相对丰度降低,厚壁菌门与拟杆菌门相对丰度比值(F/B)增加;属水平下,Stress组肉鸡盲肠中拟杆菌属(Bacteroides)和罗姆布茨菌属(Romboutsia)相对丰度降低,乳杆菌属(Lactobacillus)、肠球菌属(Enterococcus)和埃希氏菌-志贺氏菌属(Escherichia-Shigella)相对丰度升高(p<0.05,图 6j、6k、6l)。LEfSe多级物种差异判别分析显示(图 6b),在Control组中,Bacteroidota及其下属的Bacteroides、毛螺菌科(Lachnospiraceae)和梭菌纲(Clostridia)为核心优势标记物。热应激组的核心标记物为Bacillota及其下属的乳杆菌科(Lactobacillaceae)、Lactobacillus和联合乳杆菌属(Ligilactobacillus),提示慢性热应激导致肉鸡肠道微生态病理性失衡。
2.1. 慢性热应激肉鸡模型建立验证分析
2.2. 慢性热应激对肉鸡生长性能的影响
2.3. 慢性热应激对肉鸡肠道组织结构变化的影响
2.4. 肠道免疫组织化学和PAS染色结果
2.5. 肠道TUNEL染色结果
2.6. 慢性热应激对肉鸡血浆氧化应激指标的影响
2.7. 慢性热应激对肉鸡血浆炎症因子水平的影响
2.8. 慢性热应激对肉鸡肠道菌群结构的影响
-
本研究通过构建21 d慢性热应激肉鸡模型,系统探究了热应激对肉鸡生长性能、肠道形态结构、屏障功能、氧化应激与炎症反应及肠道菌群组成的影响。结果发现,慢性热应激可成功激活肉鸡下丘脑-垂体-肾上腺轴(Hypothalamus-Pituitary-Adrenal,HPA),显著抑制生长性能;破坏肠道黏膜形态,以空肠损伤最为严重;通过抑制紧密连接蛋白表达、减少杯状细胞数量及诱导肠上皮细胞凋亡,破坏肠道物理与化学屏障;诱导机体氧化应激与全身性炎症反应,同时改变肠道菌群的群落结构与多样性,引发菌群失调,最终通过多途径协同加重肠道损伤,并进一步加剧生长性能的下降,形成“氧化应激-屏障损伤-炎症-菌群失衡”的恶性循环。
研究结果显示,Stress组肉鸡血浆CORT和NE水平显著上调,且体温显著升高,证实了热应激模型的有效性,这与文献[16]结论——热应激激活下丘脑-垂体-肾上腺轴一致,CORT和NE的分泌水平可直接反映应激的强度与持续时间。文献[17]表明,在温度34 ℃条件下饲养AA肉鸡21 d,其ADG和ADFI下降。文献[18]中对21日龄肉鸡进行的7、14、21 d慢性热应激(35 ℃)实验也有同样发现。本研究显示,Stress组肉鸡的ADG和ADFI降低而F/G升高,这验证了热应激对肉鸡生长性能的抑制作用。除了ADFI下降这一直接因素外,肠道结构与功能的损伤导致的营养吸收障碍,以及机体的代谢紊乱,是生长性能受损的核心机制。多项研究表明,热应激会破坏肉鸡肠道形态结构完整性,导致V降低、C增加、V/C下降[19-21]。本研究结果显示,热应激导致肉鸡十二指肠、空肠、回肠的V降低、C增加、V/C下降,其中空肠的损伤程度最为显著。结合空肠作为肉鸡营养吸收主要部位的生理特性,以及本研究中空肠紧密连接蛋白表达量大幅下降、凋亡细胞数显著升高的结果,可推论空肠是慢性热应激损伤肉鸡肠道的关键靶部位。
肠道屏障功能的完整性是抵御病原体和毒素入侵的关键。文献[22]研究表明,热应激显著降低了肉鸡空肠Claudin-1、Occludin的表达和杯状细胞数量。文献[23]发现,热应激显著降低了空肠黏膜Claudin-1、Claudin-5、Occludin以及ZO-1 mRNA表达量。本研究显示,Stress组紧密连接蛋白Claudin-1和Occludin的表达量显著降低,杯状细胞数量减少,与前人研究结果类似,表明热应激会破坏肉鸡肠道物理屏障和化学屏障,增加肠道通透性。同时,肠上皮细胞的异常凋亡与屏障结构破坏形成的恶性循环,会持续削弱肠道的生理功能,这一发现补充了既往研究仅关注屏障蛋白表达的局限性,揭示了细胞凋亡在肠道屏障损伤中的关键作用。
正常生理状态下,机体抗氧化系统(SOD、CAT、GSH-Px等)与活性氧(Reactive Oxygen Species,ROS)的产生处于动态平衡,而热应激会导致抗氧化系统失调,ROS大量蓄积,机体出现氧化应激,ROS的蓄积可激活核因子-κB(Nuclear Factor-KappaB,NF-κB)等炎症信号通路,促炎细胞因子释放。有研究表明,热应激可以显著增加肉仔鸡肠道中IL-6和TNF-α的表达水平,增加肠道黏膜屏障的通透性和炎症细胞的浸润[24]。文献[25]研究发现,慢性热应激会显著降低肉鸡肠道的抗氧化酶活性,显著增加肠道中MDA和IL-1β、IL-6、TNF-α的含量。本研究结果显示,Stress组肉鸡血浆T-AOC水平和SOD、CAT、GSH-Px活性显著降低,MDA含量升高,同时促炎细胞因子IL-1β、IL-6、TNF-α水平上调,提示热应激确实对肉鸡的肠道抗氧化能力产生了不利影响,并证实了ROS蓄积可通过激活NF-κB通路引发全身性炎症反应,形成氧化应激-炎症反应的协同效应[26]。
肠道菌群参与营养代谢、屏障功能维持和免疫调节等多种生理过程。文献[27]研究表明,热应激可降低肉鸡盲肠微生物多样性,改变肠道微生物组成和丰度。文献[21]发现,热应激显著升高了肉鸡盲肠微生物Shannon指数,显著降低了Simpson指数,Beta多样性改变。本研究结果显示,热应激显著降低了肉鸡肠道菌群的OTU总数和α多样性指数,且Control组与Stress组的菌群结构在β多样性分析中完全分离,提示热应激改变了肠道菌群的群落结构与多样性。肉鸡盲肠中优势菌门常为厚壁菌门和拟杆菌门,厚壁菌门与拟杆菌门相对丰度比值是衡量肠道菌群群落结构的重要指标[28]。本研究发现,在门水平上,热应激导致肉鸡盲肠厚壁菌门和变形菌门丰度显著升高、拟杆菌门显著丰度降低,厚壁菌门/拟杆菌门比值升高,这与文献[29]在肉鸡热应激研究中的菌群结构变化趋势一致。文献[30]研究发现,21日龄肉鸡经历33 ℃热应激21 d后,其盲肠乳酸杆菌数量显著降低,同时大肠杆菌和梭菌数量显著增加。本研究结果显示与之一致的变化趋势,在属水平上,Stress组有益菌属(拟杆菌属、罗姆布茨菌属)丰度降低,肠球菌属及埃希氏菌-志贺氏菌属等致病菌丰度升高,其中乳杆菌属的异常增殖推测可能是菌群的代偿性反应。LEfSe分析进一步明确,Control组核心优势菌群为拟杆菌门及其下属有益菌,而Stress组核心标记物转变为厚壁菌门相关类群,证实热应激造成了菌群结构的病理性转变。综上所述,慢性热应激引发的肠道屏障损伤(物理、化学、生物屏障)与菌群失调(多样性下降、有益菌减少、潜在致病菌富集)互为因果,形成“氧化应激-屏障损伤-炎症-菌群失衡”的恶性循环。
本研究仍存在一定的局限性。第一,研究仅聚焦于肉鸡整体层面的生理生化指标与肠道组织学变化,未深入探究细胞水平和分子水平的特异性信号通路(如MAPK、Nrf2等)在热应激致肠道损伤中的调控作用,难以精准解析营养干预因子的作用靶点;第二,肠道菌群分析仅采用16S rRNA测序技术,未结合代谢组学探究菌群代谢产物的具体变化,无法全面阐释菌群失调与肠道损伤的因果关系。未来将从两个方向展开深入研究:一是结合转录组学、蛋白质组学与体外细胞模型,解析热应激致肠道损伤的关键信号通路,为靶向调控提供分子依据;二是联合菌群移植与代谢组学技术,明确肠道菌群代谢产物在热应激肠道损伤中的作用机制,探索通过益生菌干预改善肠道微生态的可行性。
-
本研究发现慢性热应激通过激活HPA轴,升高肉鸡血浆应激激素水平,降低肉鸡生长性能,破坏肉鸡肠道形态结构与物理、化学屏障功能,诱发氧化应激和炎症反应,同时造成肠道菌群微生态失衡,形成“氧化应激-屏障损伤-炎症-菌群失衡”的恶性循环。