





-
开放科学(资源服务)标志码(OSID):

-
喀斯特石漠化地区的生境脆弱性往往导致植物多样性较低[1-2],且对植物种类成分具有强烈的选择性[3]. 苔藓植物能够广泛分布在喀斯特石漠化地区,并在成土、保湿方面为其他高等植物种子的萌发和生长提供了有利条件,成为喀斯特地貌下生物多样性的重要组成部分[4-8];同时,苔藓植物的生长受小环境的影响非常显著,其功能性状能够对所处环境做出灵敏的反应[9].
当前学界对于喀斯特地区苔藓植物物种多样性、生态分布、生活型以及结皮功能与环境因子之间的关联开展了大量的研究工作:任青青[10]对喀斯特苔藓研究发现小生境异质性导致石生苔藓植物组成具有明显的差异性;Kimmerer等[11]和John等[12]对苔藓物种丰富度与小环境关系研究发现,石生苔藓物种多样性与基质岩石类型及大小、岩面的角度和方向存在密切联系;涂国章等[13]对喀斯特地区石生苔藓生态分布与环境因子之间的关系研究发现,影响石生苔藓分布的主要环境因子为坡度、土壤厚度、pH等;龙明忠等[14]对喀斯特地貌下苔藓植物的微气候效应研究发现,苔藓植物影响周围局部微气候,对微气候具有增加湿度、降低温度的作用;申家琛等[15]和程才等[16]研究阐明了苔藓结皮能够有效改善小环境,促进植被演替,增加生物多样性,利于石漠化生态系统的正向演替. 这些工作有效补充了苔藓植物与环境关系的研究,但苔藓植物功能性状与微生境因子之间的联系鲜有报道.
因此,本文选择中梁山喀斯特石漠化区域的苔藓植物群落为研究对象,对不同微生境类型下苔藓植物的部分功能性状进行探究,以期弄清苔藓植物功能性状与微生境类型及因子之间的相互关系,进而为喀斯特石漠化地区的植被恢复提供参考.
HTML
-
研究区域在重庆市中梁山(106°35′31″E,30°05′36″N). 中梁山位于重庆市北部,地处四川盆地东部,川东平行岭谷地带,地势呈北向南倾斜,起伏大,有大量的喀斯特石漠化景观. 海拔500~700 m,属亚热带湿润季风气候,雨量充沛,热量丰富;多年平均气温17.3 ℃,年平均降水量1 000~1 300 mm,降雨集中在每年4-9月,5月最多,占全年降水的70%. 地貌类型以石灰岩山地为主,属于喀斯特低山峰丛地貌,地貌类型受到地质构造和岩性的强烈控制;土壤发育的背景是三叠纪嘉陵江组的岩溶砾状白云质灰岩,土层浅薄,土被不连续,岩石裸露率高[17]. 研究区域石灰岩裸露,石缝甚多,坡上生长大量的苔藓植物,并随着微型地貌的改变呈现不同的状态.
-
研究样地大小为20 m×20 m,整块、裸露植被覆盖很低的石灰岩,坡度、坡向一致,主要植被为苔藓植物,经鉴定种类为长叶曲柄藓(Campylopus atro-virens de not). 在样地内设置31个苔藓植物群落样方,调查每个样方所在的微生境类型、微生境宽度、微生境长度、微生境深度、相对光照、土壤pH和土壤厚度等因子,同时调查苔藓植物的生物量、盖度和厚度等指标以表征苔藓植物的功能性状. 微生境类型判定参考杨瑞等[18]和罗海波等[19]的研究(表 1).
首先根据表 1判断调查样方属于何种微生境类型;然后用样方框统计样方内苔藓盖度,用铁纤和直尺测量苔藓厚度和土壤厚度[20],用照度计测量相对光照[21],用直尺测量微生境长度、宽度和深度. 每个指标测量重复5次并取平均值;最后用铲子采集样本带回实验室供进一步分析、测试.
-
用筛网和镊子将苔藓植物体和土壤基质分离,再将苔藓样本用水浸泡洗净、自然风干,然后将苔藓植物体放入烘箱内80 ℃烘72 h,样本取出称干质量. 将土壤基质与水按1∶1混合,静置1 h后用pH计测量土壤pH[22]. 记录实验结果,每个指标测量重复5次并取平均值.
-
藓类生物量的计算参照徐杰等[23]的计算公式
式中,M为藓类生物量(g/cm2),m为藓类干质量(g/cm2),C为盖度(%).
数据分析采用Excel 2019,SPSS 26软件,显著性水平设定为α=0.05. 采用one-way ANOVA分析不同微生境类型生物量的差异;用Pearson相关性系数初步分析苔藓植物功能性状和环境因子间的互相作用. 采用Canoco 5软件对苔藓功能性状和环境因子进行RDA分析(冗余分析),作图使用Excel软件.
1.1. 研究区域概况
1.2. 野外调查及采样
1.3. 样本处理和测试
1.4. 数据处理与分析
-
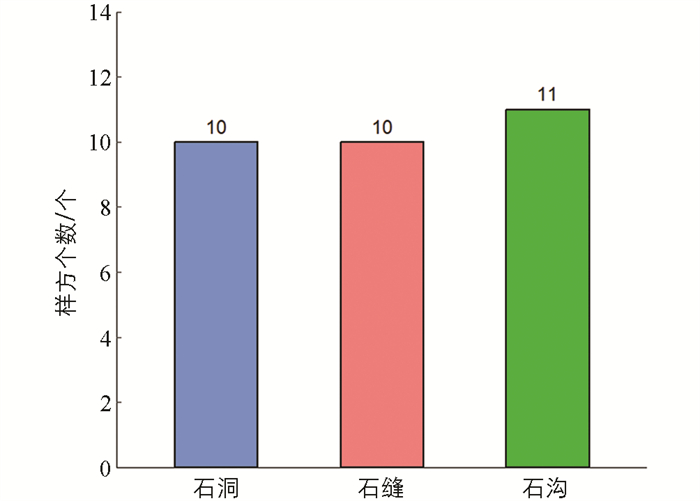
不同微生境分类如图 1,在实验区域中石缝类微生境10个,石沟类微生境11个,石洞类微生境10个,共31个样方,其中石沟类微生境数量较多,石缝和石洞类微生境数量一样,3种类型数量整体差别不大.
-
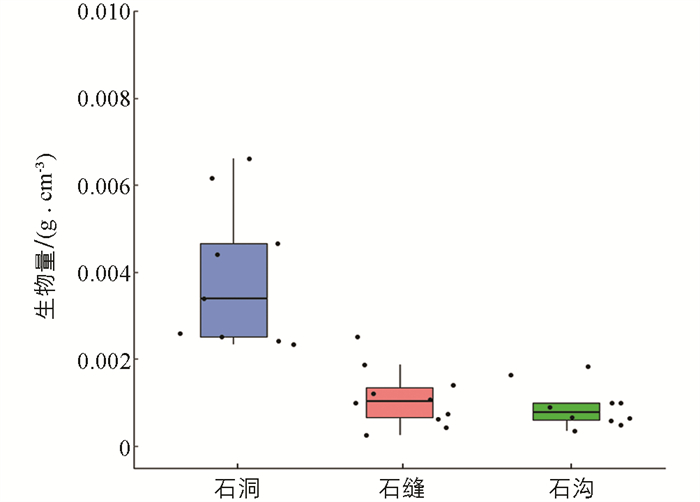
不同类型微生境苔藓群落生物量的比较如图 2. 石洞、石缝和石沟生物量平均值分别为0.003 3 g/cm2,0.001 1 g/cm2,0.000 97 g/cm2. 由不同类型微生境生物量方差齐性分析检验结果可知,Levene Statistic为5.696,p=0.008 < 0.05,非齐性. 因此对不同类型微生境生物量进行Brown-Forsythe检验和Welch检验(表 2),3种不同类型微生境苔藓植物生物量有统计学意义. 对不同类型微生境生物量进行Dunnett T3检验配对比较(表 3),显示石洞苔藓生物量相比石缝类和石沟类微生境生物量差异有统计学意义. 石洞类微生境样本生物量分布较离散,而后两者样本生物量相对集中.
-
Pearson相关性分析表明了微生境因子与苔藓植物功能性状之间的相关性. 由表 4可知,微生境宽度与苔藓盖度、苔藓厚度和生物量之间呈极显著正相关,微生境深度与苔藓盖度、苔藓厚度和生物量之间呈极显著正相关,土壤厚度与苔藓厚度和生物量之间呈极显著正相关;土壤pH与生物量和苔藓盖度之间呈极显著负相关. 微生境长度与各苔藓功能性状之间均没有显著相关性. 微生境宽度和微生境深度之间呈极显著正相关,土壤厚度与微生境宽度和微生境深度之间呈极显著正相关,土壤pH与微生境宽度呈极显著负相关,表明不同环境因子之间也会相互影响. 相对光照主要与群落上面的植被覆盖有关,与其他微生境因子和功能性状均没有显著相关性.
-
冗余分析是回归分析结合主成分分析的排序方法,能够反映响应变量与解释变量之间的相关关系[24],广泛应用于生态学的研究中[25]. 为更好地揭示微生境因子与苔藓植物功能性状之间的相互关系,本实验将苔藓盖度、生物量、苔藓厚度作为石生苔藓植物的功能性状指标,微生境因子包括微生境长度、微生境宽度、微生境深度和土壤厚度、土壤pH以及相对光照. 对两组数据进行冗余分析(图 3),黑色箭头代表微生境因子,红色箭头代表苔藓植物功能性状. 黑色连线越长,表明在环境因子中影响占比越大,反之越小. 黑色箭头与红色箭头之间的夹角,反映环境因子与功能性状的相关性. 黑色箭头之间夹角反应各环境因子之间的相关性,夹角越小,相关程度越大,若成直角则说明无相关性,夹角为钝角时,表示呈负相关.
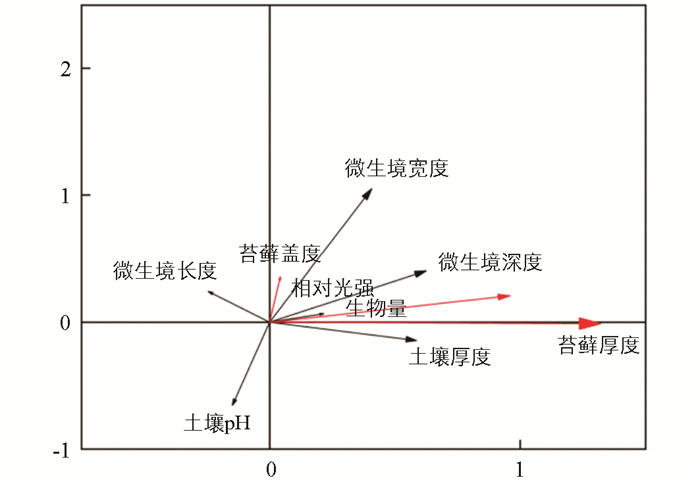
经RDA排序后可知,微生境因子对功能性状的解释量为78.87%. 第一轴和第二轴中,苔藓植物功能性状和微生境因子相关性累计百分比为77.67%和1.20%,其特征值分别为0.688 2和0.071 9,都大于0.05,达到显著性水平[26]. 微生境因子中对苔藓植物功能性状有影响的因子为微生境深度、微生境宽度、土壤厚度和相对光照. 除微生境长度和土壤pH外,各微生境因子之间均存在显著的正相关. RDA分析发现相对光强对苔藓植物功能性状也有影响,但影响较弱;微生境长度与苔藓植物功能性状没有明显关联;土壤pH与苔藓植物功能性状之间呈极显著负相关. 以上结果都与Pearson分析相吻合.
2.1. 微生境类型
2.2. 不同类型微生境苔藓群落生物量的比较
2.3. 苔藓植物功能性状与微生境因子之间的相关性分析
2.4. 苔藓功能性状与环境因子之间的冗余分析(RDA)
-
为了控制环境变量对苔藓植物功能性状影响的一致性,本研究选择在坡度、坡向和岩性基本一致的缓坡上进行. 调查微生境包括石沟类11个,石缝类10个和石洞类10个. 相关研究指出微生境的形态特征与植物生长状况密切相关[27],本研究发现石洞、石缝和石沟3种微生境类型中苔藓植物生物量有统计学意义,其中石洞类苔藓植物生物量显著多于其余二类,说明苔藓植物生物量受到微生境类型影响较大. 这与Heino等[28]和Mod等[29]的研究结果一致. 王福星等[30]和蒙文萍等[31]研究指出苔藓植物溶蚀作用对于微生境的构造也具有显著影响,苔藓植物功能性状与微生境类型存在相互作用、相互影响的关系. 苔藓植物通过吸收雨水中Ca(HCO3)2分泌酸性物质溶解岩石矿物塑造溶蚀微生境[32-33],相反微生境也可以影响苔藓植物的生长.
微生境宽度、微生境深度、土壤厚度与苔藓植物功能性状之间呈显著正相关,可能是因为微生境宽度和深度的增加拓宽了微生境体积,增大了苔藓殖居空间,促进了生物量的累积,进而影响苔藓植物其他功能性状[34]. 土壤pH与苔藓功能性状之间呈显著负相关,这与刘艳等[35]的研究结果一致,可能是因为土壤pH升高使土壤中有机质和矿物质含量降低,进而阻碍了苔藓植物对有机质和矿物质的有效吸收,抑制了苔藓植物的生长[36];也可能是土壤pH对苔藓植物并非直接抑制,而是通过影响土壤微生物区系[37],间接控制土壤矿物质转化来参与调控苔藓植物的生长. 冗余分析发现相对光强对苔藓植物功能性状也有影响,这与St.Martin等[38]的研究结果一致. 光照影响苔藓植物的光合作用,促进苔藓植物对营养元素的利用,苔藓植物生物量的增加与相对光强密不可分.
-
有关苔藓植物功能性状的研究已较多,但关于喀斯特石漠化地区微生境与苔藓植物功能性状关系的研究较少. 本研究开展了微生境对苔藓植物功能性状影响的研究,迫于实验样地条件限制,研究样本数仅有31个,数量偏少. 后续研究需加大样本数量和拓展微生境类型,并进一步探讨影响苔藓植物功能性状的主要驱动因子以及不同苔藓植物对微生境因子的响应.
由于生物量大的苔藓植物在退化生态系统修复上具有重要意义,结合本研究,在喀斯特石漠化地区进行生态修复或城市景观再造时,选择恰当的微生境和生物量大的苔藓植物,如在宽大的石洞中栽种本地优势苔藓植物,将有利于生态恢复和城市绿化的效果.
 DownLoad:
DownLoad: