





-
开放科学(资源服务)标志码(OSID):

-
叶片是植物的主要营养器官,是植物光合作用和蒸腾作用等生理代谢过程的重要场所. 叶片的发育过程是非常关键的植物形态建成过程,受生理和外界环境等多种因素的综合调控[1]. 叶片的发育过程主要包括叶片的发生、叶片的形态特征决定和叶片的极性分化3个重要过程. 叶片的极性分化包括即背-腹轴(近-远轴极性)、基-顶轴和中-侧轴3个方向的分化[2-3],其中近-远轴极性的建立能够决定叶片背腹特征的形成,形成叶片背侧的海绵组织和腹侧的栅栏组织的分化,使得叶片由辐射对称变成两侧对称,这是叶片发育的关键过程之一,是保障叶片正常进行光合作用、蒸腾作用等重要生理过程的结构基础[4-6].
多种转录调控因子相互协调表达形成复杂的调控网络,共同调控了叶片的近-远轴极性的建立过程,其中HD-ZIP基因家族(包括REVOLUTA、PHAVOLUTA和PHABULOSA基因)、ARP基因家族(ASYMMETRIC LEAVES1 (AS1)、玉米ROUGH SHEATH (RS2)及金鱼草PHAN基因)和LOB(AS2) 基因家族是主要参与叶片近轴极性建立的重要基因[7];KANADI基因家族(KAN1-4)和ARF基因家族(ARF3和ARF4) 主要参与叶片的远轴极性建成[8];小RNA包括的siRNAs、ta-siRNAs和miRNAs等也可通过调控靶基因的表达参与植物叶片的近-远轴极性的建成[9-10]. ASYMMETRIC LEAVES 1(AS1)和ASYMMETRIC LEAVES 2(AS2) 互为等位基因,编码形成的AS1与AS2蛋白均可形成转录因子复合体参与叶片近-远轴极性的建成过程. AS1属于ARP蛋白家族,是一类MYB-R2R3型转录因子,是参与DNA结合的结构蛋白[11];AS2蛋白属于植物特有的Lateral Organ Boundaries(LOB)蛋白家族,在植物中广泛存在,能够直接或间接参与植物生长发育及环境响应的转录调控过程[12-14]. AS1和AS2基因的突变会导致叶片褶皱、叶片不对称和叶边缘下卷等叶片缺陷的症状;AS1和AS2基因的超表达则会使叶片出现叶片边缘上卷和叶片收缩等近似近轴化极性的特征[15]. AS1和AS2基因家族还参与调控植物的其它发育和抗逆境胁迫等代谢调控过程,如茎的发育、维管束和根的发育等过程[16-17].
茶树是我国重要的叶用经济作物,叶片的发育及形态建成直接影响茶叶的产量和质量,所以研究茶树叶片发育调控机制对提高茶树经济价值的重要意义. 目前,茶树中AS1和AS2基因还未被关注,本研究从茶树基因组中鉴定克隆了CsAS1和CsAS2基因,对其进行了全面的生物信息学分析和表达分析,可为进一步研究CsAS1和CsAS2基因家族在茶树叶片发育过程中的重要功能提供借鉴.
HTML
-
茶树材料为2年生“福鼎大白茶” (Camellia sinensis cv. Fuding Dabaicha),种植于重庆市北碚区天生路2号西南大学校内教学实验基地. 转录组表达数据下载于“舒茶早”茶树基因组数据库TPIA(
http://tpdb.shengxin.ren/ ). 用于表达分析的茶树不同组织材料,包括成熟叶、茎、根及花等,取于“福鼎大白茶”,液氮速冻后,存储于-80 ℃超低温冰箱. 试验试剂和材料主要包括:植物RNA提取试剂盒(产于天根生化科技(北京)有限公司)、反转录酶及配套缓冲液(产于宝生物工程(大连)有限公司)、Taq DNA聚合酶、pMD18-T、DH5α大肠杆菌感受态细胞、DNA切胶回收试剂盒(产于上海源叶生物科技有限公司)和荧光定量PCR试剂超混液(产于重庆鼎国生物技术有限公司). 引物合成和基因测序由生工生物工程(上海)有限公司完成. -
按照植物RNA提取试剂盒说明书的步骤,分别提取“福鼎大白茶”茶树的根、茎材料的总RNA,采用反转录试剂盒进行mRNA的反转录,合成不同组织的cDNA,作为基因克隆及表达分析的材料. 克隆茶树AS1和AS2基因的PCR扩增体系为:94 ℃,5 min;94 ℃,30 s;50 ℃,30 s;72 ℃,2 min;35个循环后,72 ℃,5 min,DNA纯化试剂盒进行纯化回收测序和分析. 所有使用的引物详情见表 1.
-
分别从拟南芥数据库TAIR和JGI-Phytozome 13数据库(
https://phytozome-next.jgi.doe.gov/blast-search )中下载CsAS基因在9种不同植物中的核苷酸和蛋白质序列,利用TPIA数据库工具进行Blast搜索,鉴定茶树CsAS基因. 并进行了编码蛋白的分子量、等电点的蛋白性质分析;分别采用SOPM和SWISS-MODEL软件分析了CsAS1和CsAS2蛋白的结构;用Clustal X1.8,MEGA 4.0和DNAMAN等生物学软件分析了茶树CsAS蛋白的氨基酸序列,采用邻接法(Neighbor-joining)方法构建进化树,Bootstrap参数为1 000,其余参数设置为默认值;分别通过MEME、Plant-mPLoc和Plantcare软件分析了茶树CsAS蛋白的保守基序(基序数量设为5)、亚细胞定位和CsAS基因上游2 kb启动子序列中含有的顺式作用元件;利用模式植物拟南芥ATAS蛋白的相互作用网络,在数据库STRING中建立了茶树CsAS蛋白的相互作用关系网络. -
本研究从TPIA数据库中下载了茶树叶片中CsAS基因在盐、冷、干旱和MeJA等不同非生物逆境胁迫条件下的转录表达水平的数据,并利用Tbtools绘制热图. 为了验证数据库中的基因表达量,本研究还采用荧光定量PCR的方法检测CsAS基因在茶树5个组织部位(花、芽、成熟叶、茎、根)、不同发育时期的芽、不同叶位的叶片及在外源GA3、IAA处理条件下的表达量. 本研究采用水培的方式对2年生“福鼎大白茶”扦插苗进行外源激素胁迫处理,分析了CsAS基因的表达特异性,每盆24株,在盆中加入配制好的响应的营养液,加气泵通入氧气,每7天更换一次营养液,MS培养30天后进行MS/(100 mg/L)GA3和MS/(50 mg/L)IAA胁迫处理,分别在24 h和48 h后取样,液氮固样,-80 ℃保存. 采用艾德莱植物RNA提取试剂盒进行样本的总RNA的提取,并反转录成cDNA模板,用于CsAS基因表达特异性分析. 用Primer 3软件设计荧光定量引物(表 1),内标基因为茶树肌动蛋白基因Actin1. 10 μL Real-Time PCR反应体系为:SsoFast EvaGreen Supermix 5 μL,浓度为10 μmol/L的上下游引物各0.20 μL,cDNA 1 μL,ddH2O 3.5 μL,充分混匀,于Bio-Rad CFX 96实时定量PCR仪上进行扩增分析,反应程序为95 ℃,10 s;95 ℃,5 s;55 ℃,5 s,进行40个循环. 每个样品进行3次生物学重复和3次试验重复,采用2-ΔΔCT法分析结果,分别使用SPSS和ORIGIN软件进行差异显著性分析和作图.
1.1. 材料
1.2. 茶树CsAS1和CsAS2基因的克隆
1.3. 茶树CsAS1和CsAS2蛋白的生物信息学分析
1.4. 茶树AS受体蛋白基因的表达分析
-
本研究利用模式植物中AS蛋白的核苷酸和氨基酸序列信息,在茶树基因组数据中进行本地Blast搜索鉴定,共获得茶树中同源的AS1和AS2蛋白基因各1个:CsAS1和CsAS2. 如图 1,本研究分别以“福鼎大白茶”茶树茎和根为材料获得的cDNA为模板,克隆获得了CsAS1和CsAS2基因,长度分别为1 033 bp和688 bp. 如表 2,CsAS1和CsAS2基因分别编码344 aa和229 aa长度的氨基酸残基,分子量分别为39.45和25.33;蛋白的等电点分别为10.04和7.36;亚细胞定位预测分析显示2个AS蛋白均定位于细胞核,这与其它植物中的AS蛋白的定位模式一致. 如图 2,染色体定位分析结果显示2个CsAS基因分别定位于茶树的4号和10号染色体上.


-
系统进化树分析结果显示(图 3),茶树CsAS1和CsAS2蛋白与其它植物中的该类蛋白氨基酸序列具有高度的保守性,茶树CsAS1蛋白与葡萄VitAS蛋白的亲缘关系最近,而CsAS2与水稻OsAS蛋白的亲缘关系最近. 保守基序分析显示(图 4a),AS1和AS2蛋白在不同植物中均具有高度的保守性,不同植物CsAS1和CsAS2蛋白序列中保守基序的种类和数量也高度一致,但CsAS1和CsAS2蛋白之间差异比较明显,分属于不同的亚家族. 基因结构分析显示(图 4b),CsAS1基因只有1个外显子,CsAS2基因含有2个外显子和1个内含子. CsAS1蛋白属于ARP蛋白家族,CsAS2蛋白属于LOB蛋白家族. 多种植物的AS1和AS2蛋白的多序列比对结果显示(图 5),不同植物中AS1蛋白序列一致性为64.91%,AS2蛋白的一致性为53.69%;CsAS1蛋白氨基酸序列中含有2个保守的DNA结合结构域:DNA结构域Ⅰ和Ⅱ;CsAS2蛋白氨基酸序列中含有LOB家族蛋白所共有的半胱氨酸结构域CX2CX6CX3C(ZF基序)、GAS(Gly-Ala-Ser)、负责蛋白质二聚体的类亮氨酸拉链的螺旋基序LX6LX3LX6L、ICG和LZL等特殊结构域,均与该蛋白家族的功能有直接关系. 茶树的CsAS1和CsAS2蛋白均为典型的AS蛋白,推测可能与已经报道的其它植物AS蛋白具有相似生物学功能.



-
蛋白空间结构分析显示(图 6a),茶树CsAS1蛋白中α-螺旋所占的比例最大,为56.69%,其次是自由卷曲,为28.78%,延长链和β-转角所占的比例小;CsAS2蛋白中α-螺旋和自由卷曲所占的比例均较大,分别为36.68%和36.24%,延长链和β-转角所占的比例较小. 分别以已知蛋白MYB21和LOB蛋白为模板,用SWISS-MODEL软件构建的蛋白三级结构显示,CsAS1和CsAS2蛋白中均含有大量的α-螺旋和自由卷曲,但三级空间结构不同,差异较大,均与同源的拟南芥AS蛋白结构相似. 由此可推测CsAS蛋白与模式植物中的AS蛋白具有相同的生物学功能.

-
为明确CsAS1和CsAS2蛋白在茶树叶片和其它器官生长发育中的调控功能,本研究利用Real-time PCR的方法分析了CsAS1和CsAS2基因在茶树根、茎、叶、花和芽等不同组织部位的表达特异性.
如图 7a,CsAS1基因在茶树不同组织中均有表达,但在芽中的表达量远远高于其它组织;而CsAS2基因在茶树不同组织中均有表达,但在根中的表达量远远高于其它组织. 如图 7b,CsAS1和CsAS2基因在芽的不同发育时期中随着成熟度的增加均有先增加再减少的趋势. 如图 7c,CsAS1基因在叶片不同发育时期中随着成熟度的增加而减少,在幼嫩芽叶中的表达高于第一、二和三叶;CsAS2基因在叶片不同时期发育先降低,再恢复到在芽中的表达水平,在幼嫩芽叶和第三叶中的表达高于第一、二和三叶. 由此推测茶树CsAS1和CsAS2基因在茶树不同组织和叶片不同发育阶段发挥着不同的功能.

-
为分析茶树CsAS1和CsAS2基因的潜在功能,通过Plant Care软件分析了CsAS1和CsAS2基因的启动子序列. 结果如图 8所示,CsAS1和CsAS2基因启动子序列中均含有多个顺反子元件,主要包括胁迫响应元件(MYB、MYC)、光响应元件、植物激素响应元件和发育调控元件. 其中胁迫响应元件占比比较大,植物激素响应元件也有较多存在. 由以推测CsAS1和CsAS2基因可参与茶树对非生物逆境和植物激素的响应过程. 为了验证这两个基因能够参与植物激素的调控过程,本研究分析了CsAS1和CsAS2基因在外源激素胁迫条件下的表达特性,如图 8d所示,在50 mg/L IAA处理24 h和48 h后,CsAS1基因受到诱导,有较大程度的表达量增加,CsAS2基因在24 h时减少,48 h时增加;在100 mg/L GA3处理24 h和48 h后,CsAS1和CsAS2基因均受到诱导,但CsAS1增加的表达量相对比较少,CsAS2基因的表达被明显诱导,增加量接近50倍.

-
本研究利用TPIA已经公布的转录组数据分析了CsAS1和CsAS2基因在NaCl、干旱、MeJA和冷等不同非生物逆境胁迫条件下的表达分析. 如图 9所示,CsAS1基因可以被NaCl、干旱、MeJA和冷胁迫诱导表达,其中被冷驯化后表达上调,而在其余3种胁迫后表达下调;CsAS2基因仅在干旱处理72 h和NaCl处理24 h后被诱导,且表达上调. 由以上结果可以推测,茶树CsAS1和CsAS2基因可能参与茶树应对环境中盐、干旱和冷等非生物逆境胁迫的响应过程.

-
利用拟南芥同源的AtAS蛋白进行了CsAS蛋白的关联蛋白预测分析,如图 10所示,茶树CsAS1和CsAS2的蛋白与多种叶片形态建成相关的蛋白间均存在关联性,其中包含KNAT1(knotted1-like homeobox protein),KNAT2(knotted2-like homeobox protein),KAN(Homeodomain-like superfamily protein),LOB(Lateral organ boundaries domain family protein),TCP14(TEOSINTE BRANCHED、cycloidea and PCF 14),TCP15(TEOSINTE BRANCHED、cycloidea and PCF 15),AFO(Plant-specific transcription factor YABBY family protein)和HDA6(Histone deacetylase 6)等蛋白. 由此可推测,CsAS1和CsAS2蛋白是可以参与茶树叶片形态建成过程的.
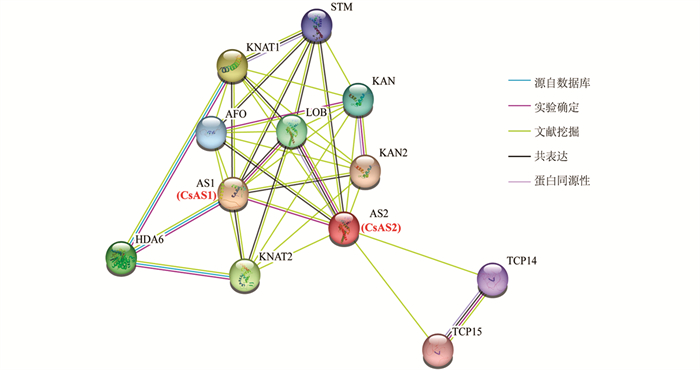
2.1. 茶树AS蛋白基因的鉴定及克隆
2.2. 茶树CsAS1和CsAS2蛋白系统进化及蛋白保守结构域分析
2.3. 茶树CsAS1和CsAS2蛋白的二、三级结构分析
2.4. 茶树CsAS1和CsAS2基因的时空表达特异性分析
2.5. 茶树CsAS1和CsAS2基因启动子片段顺式作用元件分析
2.6. 茶树CsAS1和CsAS2基因响应非生物逆境胁迫的表达特异性分析
2.7. 茶树CsAS1和CsAS2的蛋白关联蛋白分析
-
茶树是叶用经济作物,其叶片的形态建成也存在形态的多样性,例如,叶片的大小、厚度和颜色等. 其中茶树叶片的内卷和外卷等与叶片极性建立相关的现象同样也经常被观察到. 但目前茶树分子生物学研究的重点依旧在叶片的次生代谢和颜色调控上,对于叶片发育的关注度还不够. 众多转录因子参与了叶片发育和形态建成过程[18, 19]. AS蛋白已经在多种植物中被报道参与叶片的近-远轴极性建立过程,AS基因的变异或缺失会导致叶片的卷曲、外翻或者内卷等叶片去极性特征[8, 20-21]. 本研究从“福鼎大白茶”茶树中克隆获得了2个与叶片发育有着紧密相关性的基因:CsAS1和CsAS2,其分子量、等电点和编码区长度与已经报道的AS蛋白具有高度的同源性,且均定位于细胞核(表 2). 茶树CsAS1和CsAS2的氨基酸序列与水稻和葡萄等植物的该类蛋白高度相似,分别含有植物ARP蛋白的保守结构域(DNA结合结构域Ⅰ和Ⅱ)和LOB家族蛋白的保守结构域(半胱氨酸结构域CX2CX6CX3C即ZF基序,GAS,ICG和LZL区域,负责蛋白质二聚体的类亮氨酸拉链的螺旋基序LX6LX3LX6L)等特殊功能结构域. 其中,ZF基序是AS2蛋白结合DNA活性所必需的,ICG和LZL区域是AS2核定位的必要区域[22-24],说明茶树CsAS1和CsAS2蛋白具备发挥该类蛋白功能的基础序列元件. CsAS1和CsAS2基因在叶片不同发育时期中随着成熟度的增加而减少,在幼嫩芽叶中的表达量高于第一、二和三叶;CsAS1和CsAS2基因在芽的不同发育时期中随着成熟度的增加均有先增加后减少的趋势. 以上结果可说明CsAS1和CsAS2蛋白具备其它植物中报道的AS1和AS2蛋白的相似功能,可参与叶片发育中的近-远轴极性建立的调控.
AS转录因子蛋白家族还被报道广泛参与调控植物除叶片外的其它组织器官的发育和非生物逆境胁迫响应等调控过程,例如拟南芥的AS1蛋白可以负调控ABA代谢途径[25, 26],马铃薯AS2蛋白有利于提高其应对高盐、干旱、高温、低温等非生物胁迫的能力[16]. 启动子顺式作用元件和表达分析发现,茶树CsAS1和CsAS2基因的启动子序列中均含有多个逆境胁迫响应元件,茶树CsAS1基因的表达会被NaCl、MeJA和干旱胁迫抑制,可能存在负调控关系;在冷胁迫条件下,CsAS1基因的表达被上调,是正调控的关系;CsAS2基因的表达可被干旱胁迫和NaCl处理诱导. 茶树CsAS2基因在根中的表达量远高于其它组织,与玉米LOB家族蛋白RTCS和RTCL基因类似,可能能够参与侧根及卷须根的形成调控过程[27].
AS蛋白发挥生长发育调控功能是通过与其它蛋白形成复杂的网络而完成的. 茶树CsAS1和CsAS2的蛋白可与多种叶片形态建成相关的蛋白间存在关联性,通过与KNAT1、KNAT2和KAN蛋白的相互作用参与叶片的起始过程[28],和LOB蛋白家族中的其它成员相互作用,参与叶片的极性建立过程,参与TCP14、TCP15、AFO和HDA6等共同调控叶片及其它器官的发育过程[29-31]. 后续本研究团队将以CsAS1和CsAS2蛋白为中心,通过蛋白互作网络的研究,解析茶树叶片等重要组织器官的发育过程,探究CsAS1和CsAS2蛋白在茶树生命周期不同发育阶段的详细功能.
-
本论文从“福鼎大白茶”茶树茎和根中鉴定和克隆获得茶树CsAS1和CsAS2基因. 生物信息学分析显示,茶树CsAS1和CsAS2蛋白具有高度的保守性;CsAS1和CsAS2基因的表达在茶树不同组织和不同发育阶段存在时空特异性,且受到外源植物激素IAA和GA3及其它非生物逆境的影响. 根据本研究结果推断CsAS1和CsAS2基因可广泛参与茶树叶片的形态建成和非生物逆境的胁迫响应过程,为深入分析茶树CsAS1和CsAS2蛋白的功能提供了一定参考.
 DownLoad:
DownLoad: