





-
开放科学(资源服务)标识码(OSID):

-
随着矿业开发活动的深入,矿区生态环境逐渐遭遇严重破坏,土壤侵蚀、植被退化和物种多样性减少等问题亟待解决[1]。矿区生态修复的核心任务是恢复植被覆盖和生态功能,而植物的选择和生长状况直接决定了修复效果[2]。在矿区生态修复过程中,覆土厚度是影响植物生长和适应性的关键因素之一。有研究表明:过薄的覆土层可能导致土壤水分和养分供给不足,影响植物的正常生长;过厚的覆土层则可能导致土壤透气性差,影响根系的呼吸和生长[3]。覆土措施提高了植被高度和盖度,改善了渣山土壤的物理化学性质,为生态群落恢复提供基础,促进植物生长[4]。叶功能性状是植物在生存进化过程中为适应环境和提升生存能力而形成的生理或形态特征,能够反映植物物种间以及植物与生境之间的关系,体现植物与竞争者及其他生物的相互作用[5-6]。研究表明,比叶面积、叶形因子、叶干物质含量、叶氮磷含量及其计量特征是决定植物生理功能和生物地球化学循环的重要因素[7-8],而且植物叶片各性状之间存在着相互联系和相互依存的内在关系[9]。土壤是影响植物叶功能性状的重要因素,能够引起多个功能性状的协同响应[10-12],已有研究表明,土壤肥力、植物比叶面积和叶氮磷含量成正比;土壤的理化性质,如土壤水分、养分含量等,也会对植物功能性状产生显著影响[10-12]。尽管已有研究揭示了土壤特性对植物功能性状的广泛影响,但针对矿区植物叶功能性状对覆土厚度的响应机制尚未得到充分探讨。矿区土壤的特殊性,如重金属污染、养分匮乏等,可能对植物功能性状产生独特的影响,因此深入研究矿区植物叶功能性状对覆土厚度的响应机制具有重要的生态学意义。
藤本植物是指茎部无法直立生长而呈蔓生状,通常依靠攀援器官、主茎缠绕或附着于其他物体来向上攀爬的植物[13]。由于其较强的适应性和生长特性,藤本植物能够维持物种多样性与生态功能、增加绿量、缓解生态问题,成为矿区生态修复中备受关注的植物群体之一[14-15]。目前国内外学者对乔灌木、草本植物的叶功能性状与气候、资源、种内关系的研究较多,对藤本植物的研究主要集中于森林群落生态[16]、物种多样性分析[17]、林层分布规律[18]和观赏价值评价及应用[14]等方面,未对矿区中的藤本植物叶功能性状进行比较分析。对藤本植物叶功能性状的研究可以在一定程度上量化植物生长与环境的相互作用,进一步探索藤本植物的生长策略,而矿区覆土厚度的变化可能会改变藤本植物生态特性对外界环境变化的适应性策略。基于此,本研究选择6种藤本植物(五叶地锦、葛藤、常春藤、凌霄、常春油麻藤、爬山虎)为研究对象,在矿区高陡岩壁底部进行不同覆土厚度栽植原位试验,通过测量藤本植物生长情况和叶功能性状指标,探讨不同藤本植物的多个叶功能性状差异及其相关性和对不同覆土厚度的适应能力,明确6种藤本植物在不同覆土厚度下的适应性差异,筛选出耐薄土或厚土的优势物种,以期为矿区生态修复中藤本植物的选择与覆土技术的完善提供理论依据和实践参考,为矿区人工植被重建提供科学选种依据,提升生态修复效率。
HTML
-
试验地位于江西省新余市仙女湖区九龙山乡中创铁矿高陡岩壁区,地理坐标为27°36′N、114°50′E,平均海拔150.81 m,属亚热带湿润季风气候,年平均日照时间约为1 512 h,年平均气温17.5 ℃,年平均降水量1 650 mm,降雨集中在每年4-6月,6月份降雨最多,无霜期约270 d,母质为千枚岩,土壤为红壤,大部分土层厚度在10 cm以上,适宜藤本植物生长。试验地土壤前期调查数据为:土壤有机碳含量1.86 g/kg,全氮含量0.23 g/kg,全磷含量0.53 g/kg,土壤含水率22.61%,土壤pH值4.92,有效磷含量1.76 mg/kg,铵态氮含量3.42 mg/kg,硝态氮含量1.73 mg/kg。
-
2023年3月在新余市矿区高陡岩壁底部进行原位土回填覆盖试验,覆盖厚度分别为20 cm、40 cm、60 cm 3个水平,回填为宽1 m、长20 m的条带。利用6种藤本植物(五叶地锦、葛藤、常春藤、凌霄、常春油麻藤、爬山虎)对矿区高陡岩壁进行生态修复,将6种藤本植物容器苗分别移栽到3种土壤厚度中,每种厚度设置3个重复区,每个处理分别种植30株2年生容器苗,共1 620株。2024年8月进行样品采集,采集生长及结实状况良好的单株,每个处理的藤本植物选择5株生长发育良好的健壮植株,分别收集地上部分(叶、茎)、地下部分(根)生物样本,另外选取10片形状大小基本相同、叶片完全展开、无病虫害、叶边缘较完整的叶片作为样本进行表型指标测定,其余的叶片于105 ℃杀青15 min,60 ℃烘干至恒定质量,粉碎过40目筛,即为叶片养分含量待测材料。2023年2月对矿区高陡岩壁底部的覆盖土进行混合取样,带回室内进行土壤样品的肥分指标(土壤含水率、有机质、全量养分、有效氮、有效磷)分析[19]。
-
使用电子天平称取每株个体10片叶鲜质量,然后将所有叶片平铺于分辨率300 dpi的扫描仪(Epson PerfectionV 19)进行扫描,扫描的叶片图片用万深LA-S叶分析软件进行批量处理,计算叶面积、叶周长、叶长、叶宽、叶形指数、叶形状因子。将扫描后的叶片放入60 ℃烘箱48 h后取出,称其叶干质量,然后结合叶干质量和叶面积计算比叶面积和比叶质量,结合叶片干质量和鲜质量计算叶含水率、叶肉质化程度和叶干物质含量[20]。另外,分别称取适量的地上部分(叶、茎)、地下部分(根)的干样,测定其氮磷含量并计算氮磷比[21]。
-
使用Excel 2010软件和IBM SPSS statistics 26.0软件对数据进行统计汇总分析。利用单因素方差分析法比较6种藤本植物之间和不同覆土厚度下叶功能性状及氮磷养分策略差异(α=0.05);用双因素方差分析法检验藤本植被类型与覆土厚度的交互作用,并用Duncan法进行多重比较(α=0.05);采用Origin Pro 2024软件进行Pearson相关性分析和主成分分析;运用隶属函数法对不同覆土厚度下6种藤本植物叶功能性状及氮磷养分策略进行综合评价。
式中:LSI代表叶形指数;LL代表叶长;LW代表叶宽。
式中:LSF代表叶形状因子;LA代表叶面积;LP代表叶周长。
式中:SLA代表比叶面积;LA代表叶面积;LDW代表叶干质量。
式中:LSW代表比叶质量;LDW代表叶干质量;LA代表叶面积。
式中:LSD代表叶肉质化程度;LFW代表叶鲜质量;LDW代表叶干质量;LA代表叶面积。
式中:LDMC代表叶干物质含量;LDW代表叶干质量;LSFW代表叶饱和鲜质量。
式中:LMC代表叶含水率;LDW代表叶干质量;LFW代表叶鲜质量。
式中:LN(P)U代表叶氮(磷)素吸收量;LN(P)C代表叶氮(磷)含量;LB代表叶生物量。
式中:SN(P)U代表茎氮(磷)素吸收量;SN(P)C代表茎氮(磷)含量;SB代表茎生物量。
式中:RN(P)U代表根氮(磷)素吸收量;RN(P)C代表根氮(磷)含量;RB代表根生物量。
式中:TN(P)U代表总氮(磷)素吸收量。
1.1. 试验地概况
1.2. 试验设计
1.3. 叶功能性状和养分指标测定
1.4. 数据处理
-
由表 1可知,植被类型、覆土厚度及其交互作用对各项生长情况指标均存在极显著影响。五叶地锦在20 cm和60 cm覆土厚度下表现出较高的SL和SB,其中SB显著高于其他5种藤本植物,平均值分别达8.13 g和21.08 g;在60 cm覆土厚度下的SMC显著低于其他5种植物;SMC随覆土厚度的增加而增加,但RB反之。葛藤在3种覆土厚度下的RB、TB、LMC和SMC均要高于其他5种植物;在40 cm覆土厚度下的LB和RMC也显著高于其他藤本植物;在60 cm厚度下,大部分指标均优于其他藤本植物;除SL、RB及RMC外,其他生长情况指标随覆土厚度的增加而升高。常春藤在3种覆土厚度下各项生长指标均较低,其中在40 cm和60 cm覆土厚度下的SL、LB、SB、RB、TB和RMC均显著低于其他5种植物;SL随覆土厚度的增加而增加。凌霄的SL平均值在40 cm覆土厚度下是葛藤的4.52倍;RMC在60 cm覆土厚度下最高(64.12%),而LMC最低(41.70%);LB、SMC和RMC随覆土厚度的增加而增加,而RB和LMC反之。常春油麻藤在20 cm覆土厚度下LB、SMC和RMC显著高于其他藤本,在60 cm覆土厚度下LB要显著高于其他植物;LB随覆土厚度的增加而增加,而LMC反之。爬山虎在3种覆土厚度下的各项生长指标总体上要低于其他植物,其中在20 cm覆土厚度下的LB、RB和TB显著低于其他5种植物;仅在40 cm覆土厚度下的SB平均值最高(10.70 g),但LMC、SMC平均值最低;在60 cm覆土厚度下的RB平均值最低(1.58 g),显著低于其他5种植物;SL和LB随覆土厚度的增加而增加,而RMC反之。
-
由表 2可知,植被类型及其与覆土厚度的交互作用对各项叶功能性状指标均存在极显著影响,除LSF外,覆土厚度对各项叶功能性状指标均存在显著影响。五叶地锦的LA、LP、LL、LSW和LSD随覆土厚度的增加先降低后增加,但LSF、SLA和LDMC与之相反;仅LSI随覆土厚度的增加而增加。葛藤在覆土厚度20 cm下LA、LL、LW及SLA均显著高于其他5种植物,但其LSW和LDMC要显著低于其他5种植物;随覆土厚度增加,其叶片多项指标均先降低后增加,如LA、LP和SLA,而LSF、LSW和LSD先增加后降低,LL和LSI则表现为持续降低。爬山虎的LSW、LSD在20 cm和40 cm覆土厚度下均要高于其他藤本;随覆土厚度增加,LA、LP、LL、LW呈增加趋势,但LSD随覆土厚度增加而降低。常春油麻藤在40 cm覆土厚度下LA、LP、LL、LW和LSI均显著高于其他5种植物;在60 cm覆土厚度下LA、LP、LL、LW和LSI要显著高于其他5种植物;LA、LL、LW、LSF、SLA和LDMC随覆土厚度增加而增加,但LSW和LSD反之。凌霄的LSI和LSF随覆土厚度的增加而降低,但LDMC反之。
-
由表 3可知,植被类型及其与覆土厚度的交互作用对各项氮磷养分分配特征指标均存在极显著影响;除LPC外,覆土厚度对各项氮磷养分分配特征指标均存在显著影响。五叶地锦的SNC、RNC随覆土厚度的增加显著降低;葛藤的RPC随覆土厚度的增加而增加;常春藤的LNC和LN/P随覆土厚度的增加而降低;凌霄的SNC、SPC和RPC随覆土厚度的增加而增加,但SN/P和RN/P反之;常春油麻藤的SNC、RNC、SPC和RN/P随覆土厚度的增加显著提升;爬山虎的LNC和LPC随覆土厚度的增加显著提升,但RNC却与之相反。在不同覆土厚度下不同藤本植物的LN/P平均值均低于14.00 mg/g。在覆土厚度20 cm下,五叶地锦的LPC和SPC均要显著高于其他藤本植物;常春藤的LPC最低;爬山虎的RNC和RPC均要显著高于其他藤本植物;凌霄的LNC、SNC、RNC、SPC、RPC和LN/P较低。在覆土厚度40 cm下,五叶地锦的LNC和LN/P均要显著高于其他藤本植物;葛藤的SNC最高;凌霄的SNC和RPC均显著低于其他植物,但LPC最高;爬山虎的3个部位磷含量均较高。在覆土厚度60 cm下,五叶地锦的SNC、RNC、SN/P和RN/P显著低于其他藤本植物;葛藤的LNC、常春油麻藤的SNC、RNC、SPC、SN/P和RN/P以及爬山虎的LPC、RPC均显著高于其他藤本植物。
由表 4可知,覆土厚度、植被类型及其交互作用对不同覆土厚度下藤本植物各部位氮磷养分吸收量有显著影响。在覆土厚度20 cm下,五叶地锦的SNU、SPU、RPU和TPU均要显著高于其他藤本植物;葛藤和常春油麻藤的LNU较高,葛藤的RNU和TNU均要高于其他藤本植物;常春藤和爬山虎多个指标均较低,其TNU和TPU均要低于其他4种藤本植物;LPU较高的有五叶地锦、葛藤、凌霄和常春油麻藤,RPU仅五叶地锦和葛藤相对较高。在覆土厚度40 cm下,葛藤的SNU、RNU、LPU、RPU、TNU和TPU均显著高于其他植物,TNU和TPU分别是常春藤的11.83倍和12.00倍;常春藤根、茎、叶的氮磷吸收量均显著低于其他藤本植物;常春油麻藤的LNU最高(69.22 mg),是常春藤的6.22倍。在覆土厚度60 cm下,葛藤和常春油麻藤的多个部位氮磷吸收量较高,其中葛藤的LNU、RNU、RPU、TNU和TPU均显著高于其他植物;常春藤和凌霄的茎、根氮磷吸收量较低。随覆土厚度增加,凌霄和爬山虎的LNU呈增加趋势,而五叶地锦和常春藤的LNU与之相反;凌霄和爬山虎的LPU随覆土厚度增加而增加;葛藤和常春油麻藤的LNU、SNU、LPU、SPU和TPU随覆土厚度的增加而增加,常春油麻藤的RNU和TNU也表现出随之增加的趋势。
-
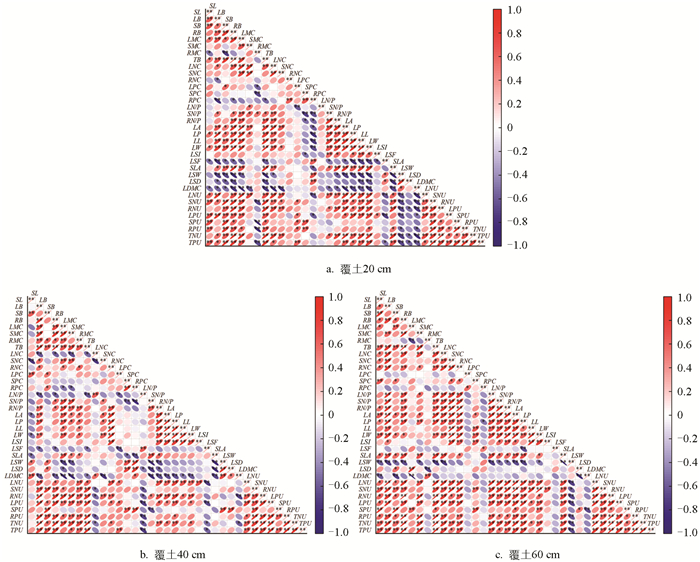
图 1是一张整合了同一覆土厚度下6种植物所有样本数据的相关性热图,计算的是在所有植物、所有样本混合在一起的情况下,叶功能性状与氮磷养分指标之间的总体相关系数。横、纵坐标均表示35个指标,右侧刻度标表示指标间相关系数的数值范围。如图 1所示,不同覆土厚度下各藤本植物的叶功能性状和氮磷养分特征的相关性各不相同。在覆土20 cm时(图 1a),SL、LB、SB、RB和LMC与TB、LP、LW、SNU、LPU和TPU呈极显著正相关,与LSF、LSW和LDMC呈显著负相关;TB、LNC与RN/P、LA、LP、LL、LW、SLA、LNU、SNU、RNU、TNU和TPU呈极显著正相关,与LSW和LDMC呈显著负相关;RPC与大部分叶片指标均呈显著负相关,仅与LSF、LSW和LSD呈极显著正相关;SN/P、RN/P、LA、LP、LL和LW与LSI、SLA、LNU、SNU、RNU和TNU呈显著正相关,而与LSD、LSW和LDMC呈显著负相关;LSW和LDMC与LNU、SNU、RNU、LPU、RPU、TNU和TPU呈显著负相关;SLA、LNU和SNU与RNU、LPU、TNU和TPU呈极显著正相关;RNU、SPU、RPU和TNU均与TPU呈极限正相关。在覆土40 cm时(图 1b),LB、LMC和RMC与TB、RN/P、LA、LP、LL、LW、SNU、RNU、LPU、RPU、TNU和TPU呈显著正相关;RNC与大部分叶片指标均呈极显著正相关;RN/P、LA、LP、LL和LW与LSI、LNU、RNU、LPU和TNU呈显著正相关,而与LDMC呈显著负相关;LNU、SNU和RNU与LPU、RPU、TNU和TPU呈极显著正相关。覆土60 cm时(图 1c),LB和RB与TB、LNC、RNC、LN/P、SN/P、RN/P、LA、LP、SLA、SNU、RNU、LPU、RPU、TNU和TPU呈极显著正相关,与LSW呈极显著负相关;TB、LNC、SNC、RNC与LN/P、SN/P、RN/P、LW、LNU、SNU、RNU、LPU、TNU和TPU呈极显著正相关;LN/P、SN/P、RN/P、LA、LP、LL、LW和SLA与大部分叶片指标均呈极显著正相关,仅与LSW呈显著负相关;LSW与大多数指标呈显著负相关;LNU、SNU、RNU、LPU、SPU、RPU、TNU和TPU与各指标之间基本呈极显著正相关。
-
对不同覆土厚度下的6种藤本植物的35个指标进行主成分分析(图 2、表 5)。由图 2可知:五叶地锦的主成分1(PC1)和主成分2(PC2)的贡献率分别为53.47%和35.65%,累计贡献率达到89.12%(图 2a);PC1轴(贡献率53.47%)正方向主要与LA、SL、LSD和LP等形态建成指标紧密相关,而负方向则与LDMC、LSF、SLA等保守型性状关联,这表明PC1轴代表了该物种在“快速扩张”与“保守投资”之间的主要权衡策略;葛藤的主成分累计贡献率为82.77%(图 2b),PC1轴(贡献率54.63%)正方向聚集了SB、SL、RPU和LNU等指标,表明PC1轴主导了其生物量分配与养分吸收的综合策略;常春藤的主成分累计贡献率为78.29%(图 2c),其PC1轴(贡献率53.56%)正方向与LA、SB、SNU和SPU等指标紧密相关,而LDMC、LNC、LNU等则位于负方向,这反映了常春藤的生长策略集中于光合器官构建与养分获取效率的协同变化;凌霄的主成分累计贡献率达86.18%(图 2d),PC1轴(贡献率54.13%)正方向与TB、SB、LMC和RB等指标关联,负方向则与LDMC、RMC、RPC、LB、SLA等相关,此格局揭示了凌霄在响应环境时,其生物量积累、水分状况与叶片构建成本之间存在紧密的协同与权衡关系;常春油麻藤的主成分累计贡献率最高(90.01%),PC1轴贡献率达66.60%(图 2e),PC1轴正方向强烈关联LPU、RNU、SN/P和TPU指标,说明磷养分的高效利用与体内化学计量平衡是该物种最核心的适应策略;爬山虎的PC1与PC2的贡献率较为均衡,累计达87.31%(图 2f),PC1轴(贡献率47.03%)正方向与LP、SB、SNU和LB等相关,而负方向与RNC、SN/P等相关,这表明爬山虎的策略权衡同时涉及形态建成和体内养分的分配与循环。
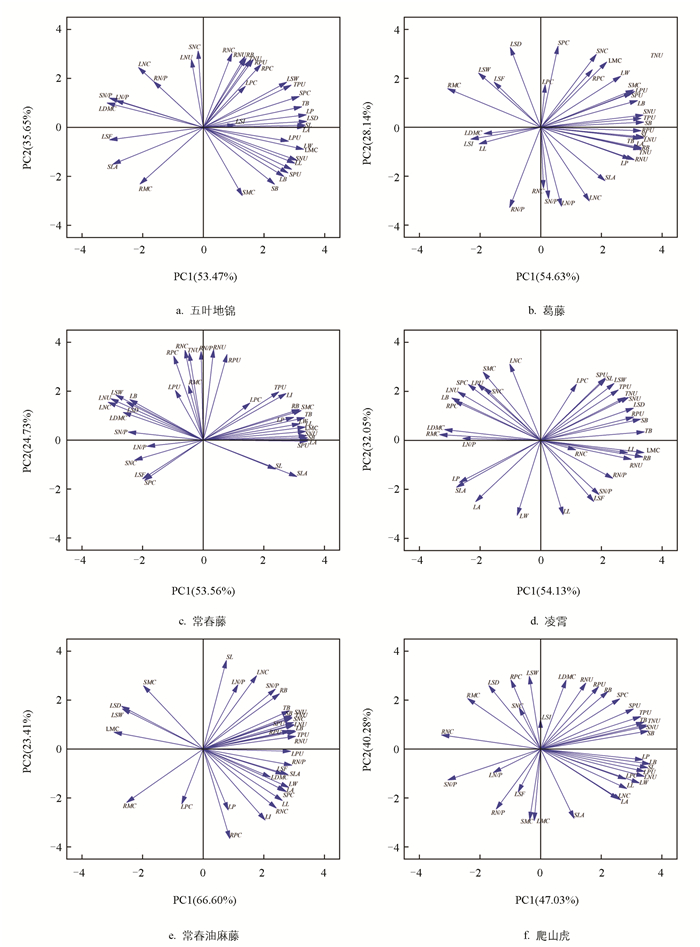
藤本植物在不同覆土厚度下2个主成分的得分如表 5所示。覆土厚度20 cm时,常春油麻藤和爬山虎在PC1负轴得分最高,葛藤和凌霄在PC2负轴得分最高,五叶地锦在PC2正轴的得分最高,说明在覆土厚度20 cm下五叶地锦长势较好,常春油麻藤、爬山虎、葛藤和凌霄长势较差;覆土厚度40 cm时,五叶地锦在PC1负轴得分最高,常春油麻藤在PC2负轴的得分最高,葛藤、凌霄和爬山虎在PC2正轴的得分最高,说明在覆土厚度40 cm下五叶地锦和常春油麻藤长势较弱,葛藤、凌霄和爬山虎长势较好;覆土厚度60 cm时,葛藤、常春藤和常春油麻藤在PC1正轴的得分最高,五叶地锦和爬山虎在PC2负轴的得分最高,说明在覆土厚度60 cm下葛藤、常春藤和常春油麻藤长势较好,但五叶地锦和爬山虎长势较弱。葛藤和常春油麻藤的3个覆土厚度的样本点沿PC1轴呈现梯度分布,揭示了厚度变化是驱动其策略转变的关键因子。其他藤本植物在不同覆土厚度下的点位分布较分散,显示出较高的表型可塑性。
利用相关公式:F(综合得分)=F1(主成分得分1)×PC1方差贡献率/累计贡献率+F2(主成分得分2)×PC2方差贡献率/累计贡献率,计算出不同覆土厚度下各藤本植物的叶功能性状和氮磷养分特征综合得分,并依据综合得分对其进行排序(表 6),发现常春油麻藤在60 cm覆土厚度下表现最佳,综合得分最高(1.05),排名第一;凌霄在40 cm覆土厚度下表现较好,综合得分0.83,排名第四;葛藤在60 cm覆土厚度下综合得分0.81,排名第五;五叶地锦在20 cm覆土厚度下表现较好,综合得分0.79,排名第六;爬山虎在40 cm覆土厚度下表现较好,综合得分0.91,排名第二,但在60 cm覆土厚度下表现较差,综合得分-0.30,排名第十一。
2.1. 藤本植物在不同覆土厚度处理下的生长情况
2.2. 藤本植物在不同覆土厚度处理下叶功能性状的差异
2.3. 藤本植物在不同覆土厚度处理下氮磷养分分配特征和吸收量的差异
2.4. 藤本植物叶功能性状特征与氮磷养分特征间的相关性分析
2.5. 藤本植物叶功能性状及氮磷养分特征综合评价
-
环境变化会引发植物一系列行为变化,其中形态和生物量的变化是评估植物对环境适应性的重要指标[22]。在异质生境中,藤本植物通过调节生物量对不同器官的投资来适应环境变化,以确保器官生长发育,维持生存繁衍能力[23]。本研究发现,五叶地锦和凌霄的叶生物量随覆土厚度的增加而增加,但根生物量反之,说明五叶地锦和凌霄协调生物量资源分配以适应外界环境变化,实现优化觅食行为。这可能是因为在薄土层的环境下,五叶地锦和凌霄会增加根系生物量,以便扩大根系的吸收范围来获取更多的养分;相反,在厚土层中土壤养分更充足,五叶地锦和凌霄会减少对根系的投资,转而增加叶片的生物量用于提高光合作用,从而优化生长。此外,葛藤和常春油麻藤的叶生物量随覆土厚度的增加而增加,该结果与文献[3]研究高寒区弃渣场植物地上生物量随着覆土厚度的增加而增加这一结果一致。由于环境资源有限,当植物增加某一器官的生物量投资时,必然会减少对其他器官的生物量投入[24],而有限的环境资源导致植物各器官之间存在竞争,许多情况下植物会采用茎沉默策略以实现整体利益最大化[25-26]。本研究中常春藤的茎生物量在覆土厚度20 cm和40 cm下显著低于叶生物量,但在覆土厚度60 cm下反之,在一定程度上说明当土壤资源减少时,常春藤会牺牲茎的投资来满足叶投资的稳定。五叶地锦和常春油麻藤在20 cm覆土厚度下茎生长迅速、攀岩能力强,更有利于矿区生态修复。凌霄在60 cm覆土厚度下根含水率最高,反映其对深层土壤水分利用能力较强。葛藤各部位生物量在3种覆土厚度处理下均高于其他藤本植物,而常春藤和爬山虎在3种覆土厚度下各项生长指标均较低,表明葛藤相较于常春藤和爬山虎更适合在该矿区生长。大多数植物叶片含水率占组织鲜质量的65%~90%[8],而本研究中凌霄在3种覆土厚度下的叶含水率均在65%以下,且其成熟叶片一般呈纸质化,说明凌霄并不适于干旱地区种植。
-
叶经济谱系能够协同并量化一系列的叶功能性状指标,反映植物在叶片结构成本与资源回报时间之间的权衡[27]。根据叶经济谱系可将植物分为“快投资-回报”型(资源获取型)和“慢投资-回报”型(资源保守型)植物,前者通常具有较高的比叶面积和氮含量,而后者则往往具有较高的比叶质量和叶干物质含量[28]。本研究中葛藤在覆土厚度20 cm下的叶面积、叶片长、叶片宽及比叶面积总体上高于其他5种植物,但其比叶质量和叶干物质含量总体上低于其他藤本植物,说明葛藤生长策略可能是通过较大、较薄的叶片和较低的干物质含量来快速扩大叶面积和提高光合效率,适应覆盖土层较厚的环境,属于“快投资-回报”型植物,该结果与文献[29]的研究一致。爬山虎在20 cm和40 cm覆土厚度下的叶面积、叶周长、叶片长和叶片宽总体上低于其他5种植物,但其比叶质量、叶肉质化程度和叶干物质含量总体上高于其他藤本,表明爬山虎在20 cm和40 cm覆土厚度下属于资源保守型植物,其对资源和养分的利用效率高,能将更多的养分投入叶片内部组织构建,对环境有更强的耐受性、抗逆性,但同时也降低了其光合作用能力[5]。叶片的寿命与比叶质量呈显著正相关关系[30],因此葛藤在覆土厚度20 cm和60 cm下叶寿命相对较短,而爬山虎在覆土厚度20 cm和40 cm下叶寿命较长。本研究中常春藤叶面积普遍较小,但叶肉质化程度与叶干物质含量较高,符合“慢投资-回报”型植物的特征[28],说明常春藤是资源保守型植物。
植物功能性状的相对稳定性和对外界环境变化的可塑性,使其能够通过功能性状的变化反映植物在不同层面(从器官、物种、群落到生态系统)适应环境的策略和机制[31]。本研究表明不同藤本植物的叶功能性状对覆土条件的响应具有物种特异性及环境依赖性。其中,五叶地锦叶形状指数随覆土厚度的增加而增加,而葛藤与之相反,表明随覆土厚度的增加五叶地锦和葛藤会优先改变叶形来优化对水分和光照的利用。凌霄的叶形指数和叶形状因子随覆土厚度的增加而降低,但叶干物质含量反之,这些变化表明凌霄在更厚的覆土下采用了更加保守的资源配置策略,减少了对叶片形态的投资,减少不必要的能量消耗,因此可以判断凌霄属于“慢投资-回报”型植物。常春油麻藤的叶面积、叶片长、叶片宽、叶形指数、叶形状因子、比叶面积和叶干物质含量总体上随覆土厚度增加而增加,但比叶质量和叶肉质化程度反之,这些特征表明常春油麻藤在不同覆土厚度下表现出更多的“快投资-回报”型特征,这是因为覆土厚度增加导致资源供给充足,从而降低了常春油麻藤对高代谢活力的需求,促使其减少叶片的光合作用投资,增加叶干质量投资的回报,并通过缩短叶寿命来优化资源使用[32]。爬山虎的叶面积、叶周长、叶片长和叶片宽随覆土厚度增加总体上呈增加趋势,但叶肉质化程度随覆土厚度增加而降低,这些特征表明爬山虎在覆土厚度较大时表现出更高的资源获取能力,同时叶肉质化程度较低意味着其生长方式更加注重快速的资源获取而非长期的资源保守,这是因为随着覆土厚度的增加,爬山虎并不再依赖较厚的叶肉来应对水分和养分压力,节省了用于增加叶片面积和比叶面积的资源,提高了光合作用效率[33]。这些适应机制帮助藤本植物在变化的环境中维持生长和繁殖的竞争力。
-
氮和磷是植物体重要的组成成分,决定了植物的光合作用、生长、繁殖和其他生理过程,被用来评估植物的生长状况是否受到氮元素或磷元素供应的限制[34-35]。6种藤本植物间的养分分配特征和吸收量存在显著差异,这与植物自身特性有关,说明不同的藤本植物对资源的利用率以及生存策略存在一定差异。文献[36]分析了植物各个器官多种功能元素的异速分配规律,发现这些功能元素在不同器官的分配具有一定的保守性。本研究发现五叶地锦在覆土厚度60 cm下茎氮含量和氮磷比相较于覆土厚度20 cm和40 cm显著降低,与其生长策略和养分分配机制有关。根氮磷比显著高于其他植物,表明其根系优先吸收氮源。随着覆土厚度增加,五叶地锦的茎、根氮磷含量提升,茎氮磷比下降,植物会根据不同部位的生理需求来调节氮磷的分配,通常植物根部用于吸收和储存养分,茎部则更多地参与传输和支持功能,而磷元素支持其细胞分裂和组织扩展[37],因此在土壤中氮磷供给增多时,五叶地锦倾向于分配更多的养分用于茎的生长和结构支持,从而使得茎部的氮含量减少,导致氮磷比下降。葛藤和常春油麻藤在覆土厚度60 cm下根氮含量高于其他植物,这是因为葛藤和常春油麻藤是豆科植物,其根系具备较强的氮吸收能力,能有效地从土壤中获取更多的氮[38],导致其叶和茎中的氮含量显著高于其他植物。随着覆土厚度的增加,土壤中养分的供应量增加,磷在土壤中相对较难移动和被吸收,促进了葛藤根部和茎部的养分积累,尤其是磷元素在根部的富集。爬山虎的叶氮磷含量在一定覆土厚度下显著增加,但其根氮磷含量相反,这是由于爬山虎在生长过程中,叶片作为光合作用的主要部位优先吸收氮磷以支持蛋白质合成和能量储存,导致叶部氮磷含量随土壤养分浓度增加而显著提高,而根部的氮磷含量相对下降,是因为爬山虎在养分分配上倾向于优先供给叶片,促进其快速生长和光合作用。
文献[34]的研究表明,叶片氮磷比大于16时,植物生长受到磷元素的限制;氮磷比小于14时,植物生长受到氮元素的限制;氮磷比介于14~16时,植物生长受氮、磷元素共同限制。本研究中,不同覆土厚度下,6种藤本植物的叶氮磷比均低于14,说明本试验区藤本植物生长受到氮的限制。本研究发现,在覆土20 cm覆土厚度下,五叶地锦的氮磷吸收量显著高于其他藤本植物,尤其是在茎和根上,葛藤和常春油麻藤的叶氮吸收量较高;在60 cm覆土厚度下,葛藤和常春油麻藤的多数部位氮磷吸收量较高。表明五叶地锦在较薄土层中展现出更强的适应性,能更好地利用土壤中的养分,而葛藤和常春油麻藤能够适应较深的土层,并且其根系能够在较深的土壤中寻找更多的养分,这有助于提高其氮磷吸收量。凌霄和爬山虎的叶氮素吸收量随着覆土厚度的增加呈上升趋势,而五叶地锦和常春藤的氮素吸收量则呈下降趋势,表明凌霄和爬山虎更适应较肥沃的土壤环境,能更好地利用土壤中的氮,而五叶地锦和常春藤在较贫瘠或养分相对较少的环境中生长较好,其对氮的需求较低。总而言之,植被类型与覆土厚度显著影响藤本植物氮磷养分分配和吸收特征,植被类型的生理特性和土壤养分的可利用性是导致这些差异的关键原因。
-
从藤本植物叶功能性状与养分特征间相关性结果可以看出,不同覆土厚度下各藤本植物的叶功能性状和氮磷养分性状的相关性各不相同。在覆土20 cm和40 cm时,叶含水率与总生物量、叶周长、叶片宽、叶磷素吸收量和总磷吸收量呈极显著正相关,说明藤本植物在面对养分贫瘠的外界环境时叶片含水率降低,更倾向于构建防御组织以抵抗外界环境胁迫[5],但也减少了植物对磷养分的吸收量。本研究发现比叶质量和叶干物质含量与叶面积、叶周长、叶片长、叶片宽、叶氮磷比、根氮磷比、叶氮含量、叶氮素吸收量、茎氮素吸收量、根氮素吸收量、叶磷素吸收量、根磷素吸收量、总氮吸收量和总磷吸收量呈显著负相关,说明高氮磷吸收量的植物往往表现为较薄的叶片,注重快速生长。这是因为藤本植物更多地将氮、磷等养分用于支持快速的生长而非增加叶片的结构密度。在覆土40 cm时,叶面积、叶周长、叶片长和叶片宽与叶氮素吸收量、根氮素吸收量、叶磷素吸收量和总氮吸收量呈显著正相关,表明了氮和磷元素在叶片发育中的重要作用。覆土60 cm时,叶生物量和根生物量与叶氮含量、根氮含量、叶氮磷比、茎氮磷比、根氮磷比、叶面积、叶周长、比叶面积、叶氮素吸收量、根氮素吸收量、叶磷素吸收量、根磷素吸收量、总氮吸收量和总磷吸收量呈极显著正相关,这是因为养分充足供应能够促进植物的生长和光合作用。然而,比叶质量与生物量之间的负相关关系表明植物在养分充足时倾向于增加叶片和根系的生长而非单纯增加干物质的积累。
植物不同器官在养分循环中扮演不同角色,各器官对养分元素的利用具有一定的相关性[39]。本研究中不同覆土厚度下的叶氮素吸收量和茎氮素吸收量与根氮素吸收量、叶磷素吸收量、总氮吸收量和总磷吸收量呈极显著正相关;总氮吸收量与总磷吸收量呈极显著正相关。说明根氮素吸收量的增加通常会带动茎、叶等其他部位氮素的增加。植物为了维持平衡和促进健康生长,氮和磷的吸收需求相互依赖,磷不仅在能量传递和细胞分裂中扮演重要角色,也参与光合作用的调节[37],因此磷的吸收能够促进叶片和茎部的氮素吸收,尤其在植物生长旺盛时期,叶片和茎部对于氮磷的需求往往是同步增加的,该结果与文献[40]的研究结果一致。通过对不同覆土厚度下各藤本植物的叶功能性状和氮磷养分特征进行主成分分析和综合得分可知,在覆土20 cm时,五叶地锦生长状况最佳,爬山虎和凌霄在覆土40 cm时生长优势较高,葛藤、常春藤和常春油麻藤在覆土60 cm时的综合得分最高,表现出较好的适应性和生长优势。
-
覆土厚度对藤本植物的生长和适应性有显著影响。不同藤本植物种类表现出不同的适应策略,如葛藤和常春油麻藤为资源获取型,而爬山虎和常春藤则为资源保守型。此外,不同覆土厚度下的6种藤本植物叶氮磷比数据表明该试验地栽植的藤本植物生长受到氮限制。在薄土层,五叶地锦具有较强的生长优势,葛藤、爬山虎、常春藤、凌霄和常春油麻藤适合栽植在较深土层中。总而言之,藤本植物通过调整生物量分配、叶片形态和养分吸收策略来适应不同覆土厚度以提升生长和竞争能力。
 DownLoad:
DownLoad: