





-
开放科学(资源服务)标识码(OSID):

-
在日益严峻的全球气候变化背景下,减少温室气体排放、降低碳排放强度、增强碳汇能力已被普遍认为是实现可持续发展和应对气候变化的关键策略[1-2]。中国作为世界碳排放大国,正积极采取措施实现双碳目标[2-3]。森林作为陆地生态系统的主体,发挥着“压舱石”和“稳定器”的重要作用,在全球碳循环和碳中和进程中具有重要作用[3-5]。森林通过光合作用将大气中的CO2转化为有机物质,并将其固持在植被各部位(树干、枝叶和根系)和土壤两大碳库中[5],从而减缓全球气候变暖趋势[6]。森林作为直接有效的碳吸收和储存载体,具有较高的生产力,其每年固碳量约占陆地植被总固碳量的4/5,同时约有85%的陆地生物量集中于森林植被中[7-8]。因而,探究乔木林的生物量及其分配特征,有利于综合评估森林生态系统固碳潜力并维护生态系统平衡[9]。
目前,乔木生物量的测定主要采用以下几种方法:实测法(涵盖皆伐法和标准木法)、相对生长法以及生物量-蓄积量模型估算法等[10-11]。针对特殊情况,研究者还发展出若干衍生方法,例如当面临无法直接采样的濒危物种时,可通过非破坏性方式间接估算(或通过测量枝干体积和密度推算[12],或借鉴其他地区同种或近缘物种的相对生长方程进行计算),既避免人为干扰,又能有效获取生物量。国内外众多学者已从不同研究视角出发,采用多种方法体系,对乔木林生物量和生产力开展多尺度系统研究。Zhang等[13]利用生物量扩展因子法(Biomass Expansion Factor Method,BEFM)计算江西省碳储量由159.1 Tg增加至276.1 Tg;任云卯等[11]采用森林蓄积量扩展法测算北京市森林碳储量为1 934.59万t,碳密度为32.35 t·hm-2;刘领等[14]应用生物量换算因子连续函数法(Biomass Conversion Factor Continuous Function Method,BCFCFM)和平均生物量法估算河南省1998-2013年森林植被碳储量及碳密度变化。我国大多数学者依托森林资源清查数据,利用生物量法或蓄积量法测算森林碳储量,然而针对同一区域同时采用不同方法进行生物量和碳储量估算的对比研究仍较少[15]。重庆市地形复杂、气候多样,乔木资源分布广泛且不均,在估算方法、数据来源及地域差异等多种因素共同作用下,其森林碳储量估算仍存在一定不确定性[16]。因此,选取生物量回归方程法(Biomass Regression Equation Method,BREM)、生物量换算因子连续函数法(BCFCFM)、生物量扩展因子法(BEFM)和可变扩展因子法(Variable Expansion Factor Method,VEFM)4种方法估算重庆市各区县乔木林生物量,旨在分析不同方法在同一区域、同一树种条件下的计算差异,并筛选出更适用于重庆市的生物量和碳储量估算方法,以期为提高重庆市森林碳储量估算的准确性和可靠性提供参考。
HTML
-
重庆市地处西南丘陵山区(28°10′-32°13′N,105°17′-110°11′E),全市面积824万hm2,地势总体由南北两侧向长江河谷逐级降低,以山地、丘陵地貌为主[5];作为典型的亚热带湿润季风气候区,重庆具有冬暖夏热、无霜期较长等特点,全年日照时数仅1 000~1 400 h,是我国日照最少的地区之一[17-18];光照、温度、水源在同一季节内协同作用,气候资源丰富[15, 18];年均气温16~18 ℃,年均降水量充沛,绝大部分地区降水量为1 000~1 350 mm[19-20]。因长江自西南向东北贯穿全境,重庆市拥有丰富的水资源。
-
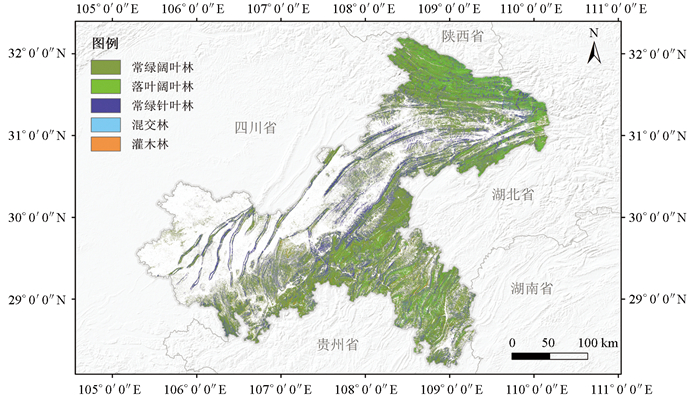
重庆市土地肥沃,水系发达,植物资源种类繁多,拥有丰富的自然资源和优越的生态环境[21-22],见图 1。自然植被类型属亚热带常绿阔叶林区,第九次全国森林资源清查期内,全市林业用地面积421.71万hm2,森林面积占林业用地总面积的84.71%,森林覆盖率达到43.11%。森林植被资源丰富,主要由亚热带常绿阔叶林、落叶阔叶林和常绿针叶林组成。针叶树种以马尾松(Pinus massoniana Lamb.)、杉木(Cunninghamia lanceolata (Lamb.) Hook.)、柏树(Cupressus funebris Endl.)为代表[21],阔叶树种包括桉树(Eucalyptus spp.)、麻栎(Quercus acutissima Carruth.)、杨树(Populus L.)、香樟(Cinnamomum camphora (L.) J. Presl)、黄葛树(Ficus virens Aiton)等[23]。
-
DEM数据来源于地理空间数据云,分辨率为30 m×30 m,数据处理环境为ArcGIS平台,投影坐标系统统一为WGS 1984,经预处理后得到高程、坡度和坡向。结合重庆市2022年森林资源一张图数据,分别提取出桉树、柏树、栎树(Quercus spp.)、杉木、马尾松、混交、其他针叶、软阔、硬阔共9类乔木林优势树种(组)。按照国家林业局森林资源调查项目规定,对森林资源调查数据进行整理,总计获得5 506 871个样本数据,并根据地类和森林类型剔除误分样本及非森林类型样本,最终共筛选提取出符合要求的1 731 412个乔木林样本数据,对应乔木林面积355.56万hm2,共包括410 571.91万株树木,总蓄积量达22 327.28万m3。各优势树种(组)样本数据具体信息见表 1。每个样本均详细记录了立地因子(坡度、坡向、海拔、土层厚度等)和林分因子(胸径、树高、郁闭度、林龄等)。
-
碳储量常用研究手段包括实地调查、遥感技术以及数学模型3类[23]。生物量-蓄积量模型多用于大尺度林分生物量估算,其原理是基于树木树干生物量、材积与其他器官间存在的强相关性,通过特定参数实现生物量数据的转换[23-26]。本研究基于森林资源数据进行估算,相较于其他方法可获得较为精确的结果。以下为本研究涉及的乔木林碳储量估算方法:
1) 生物量回归方程法
生物量回归方程法充分利用调查数据中获取的胸径、树高等较易获取的数据,在适用范围内估算误差较小,准确性较高[27]。计算公式如下:
式中:Y为乔木林地上部分森林植被碳储量;i为组成林分的树种;Ai为第i类树种的面积;D为胸径(cm);H为树高(m);a、b、c为随森林类型或气候带变化的常数;CFi为树种i的含碳率。乔木林总碳储量为各树种生物量乘以相应含碳率后的总和。
2) 生物量换算因子连续函数法
该方法亦称生物量换算因子法,假设生物量与蓄积量之间的转换关系,从而较为简便地推算大尺度森林生物量[6, 28]。由于本研究各方法采用的基础数据来源一致,因此可减少因数据来源差异造成的比较偏差;但仍需注意胸径小于5 cm的幼树未计入可能带来的低估影响。计算公式如下:
式中:Vi为第i树种单位面积蓄积量(m3·hm-2);a、b为随森林类型或气候带变化的常数。
3) 生物量扩展因子法
该方法基于树种生物量与蓄积量、基本木材密度、生物量扩展因子和根茎比之间的关系建立模型[29]。计算公式如下:
式中:SVDi为第i类树种的基本木材密度(t.d.m·m-3);BEFi为第i类树种的生物量扩展因子(无量纲);Ri为第i类树种根茎比,即地下生物量与地上生物量之比。
4) 可变扩展因子法
考虑到不同林龄对生物量的影响,在BEFM基础上利用可变的BEF和R值分别计算不同龄组各树种生物量,即为VEFM[29]。不同林龄对应的生物量扩展因子与根茎比存在差异,但变化趋势因树种而异。表 2给出了不同树种按龄组划分的生物量扩展因子BEF与根茎比R值。
-
1) 含碳系数
森林碳储量通常由生物量乘以相应含碳系数估算得到。目前国内外关于碳储量的研究通常采用国际普遍使用的乔木含碳率0.5 g C·g-1来估算,在大尺度碳储量估算中该方法能显著降低工作难度[5, 8, 22]。本研究依据国家市场监督管理总局和国家标准化管理委员会发布的《主要树种立木生物量模型与碳计量参数》 (GB/T 43648—2024)及前人研究成果,确定各优势树种(组)的含碳率[10, 11, 14, 28](表 3)。森林碳密度计算公式如下:
式中:ρ为碳密度(Mg·hm-2);Ytotal为总碳储量;Stotal为总面积。本研究中的森林碳储量仅指基于树木活体生物量估算的碳储量,不包括森林生态系统中其他碳库组分,如枯木、草本植物层、凋落物层以及土壤层等所含碳储量。
2) 其他参数
BREM中D、H数据可从森林资源清查数据中直接获取,其他系数在参考《主要树种立木生物量模型与碳计量参数》 (GB/T 43648—2024)的基础上,再结合其他学者的相关成果设定[7, 14, 28];BEFM参数依据2018年国家林业和草原局发布的《森林生态系统碳储量计量指南》 (LY/T 2988—2018)设定;VEFM转换参数详见表 2。
1.1. 研究区概况
1.1.1. 自然概况
1.1.2. 森林资源概况
1.2. 数据获取
1.3. 研究方法
1.3.1. 碳储量估算
1.3.2. 碳储量估算参数设定
-
目前针对乔木林碳储量计算结果的精度评估尚未建立统一的标准体系,因此在比较不同模型的计算精度时,难以客观判定何种方法更具准确性和可靠性。为此,从计算过程、方法特点、模型可验证性、可重复性及稳定性等方面对4种碳储量估算方法进行系统比较[27-28]。
-
采用相对误差分析方法对比结果(图 2),以平均值作为基准计算相对误差,比较确定最适用于计算重庆市森林碳储量的方法。结果表明:相对误差由大到小依次为BREM(27.17%)、BCFCFM(22.18%)、BEFM(7.51%)、VEFM(4.48%);不同优势树种组中相对误差BREM多出现较大的负误差,BCFCFM多出现较大正误差,而BEFM(-11.36%~28.68%)与VEFM(-11.46%~21.12%)误差居中,且大多数生物量和碳储量的误差幅度均小于BREM(-64.44%~-19.82%)与BCFCFM(-21.82%~65.86%);此外,碳密度的误差幅度由于面积这一变量的影响有所降低:最大误差幅度的绝对值由大到小依次为换算因子连续函数法(66.80%)、回归方程法(57.23%)、扩展因子法(39.16%)、可变扩展因子法(27.52%)。
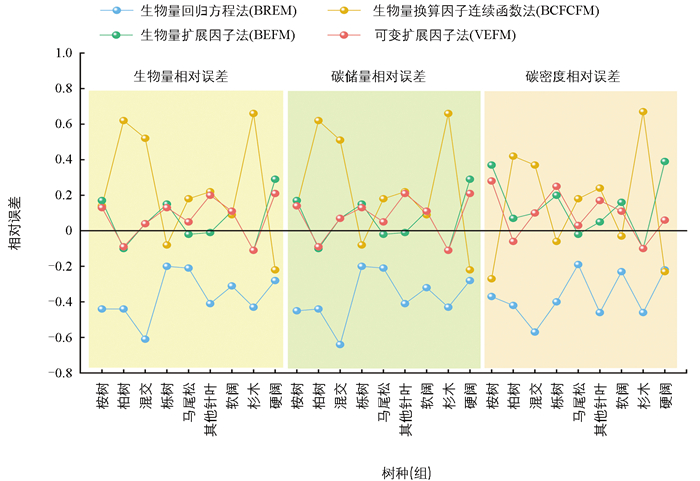
总体来看,对于马尾松、栎树、软阔而言,4种模型误差幅度均较小,表现较为稳定;而除上述3类树种外,BCFCFM和BREM在其余树种中的误差幅度相对较大,表明其稳定性较差;同时,柏树、杉木的最大误差值分布在BCFCFM中,硬阔类在BEFM中误差幅度最大;各树种误差幅度最小值范围为0.01~0.21,且桉树、柏树、混交、杉木和硬阔的相对误差幅度最小值均稳定分布在VEFM中,马尾松、其他针叶的最小误差幅度位于BEFM中,而栎树和软阔在BCFCFM中的模拟效果最为稳定。
-
4种计算方法得出的结果不同,主要源于所采用的模型与假设存在差异,不同数据来源、方法考虑因素以及模型参数设置等均会对结果产生影响。以重庆市分布最为广泛且相对误差对比最明显的马尾松为例(表 4),分析4种模型的计算过程及各模型的优缺点。
-
生物量回归方程法充分利用森林资源数据中易获取的胸径、树高等因子,构建单位面积生物量的估算模型,并在此基础上推算区域尺度生物量[19, 30-31]。以马尾松为例,模型中的估计参数a、b、c分别为0.092、2.028、0.498,计算结果显示马尾松总生物量为94.85 Tg,总碳储量为49.48 Tg。但该方法也有一定局限性,例如胸径大小主要反映林龄,树高在一定程度上体现立地质量,而原始数据测算精度和适用范围均会直接影响结果的准确性,即该模型的适用性存在一定局限。对于总生物量而言,BREM方法的计算结果偏小。
-
马尾松参数a(基础转换参数,仅与蓄积量有关,与林龄无关[27, 31])、b(调节转换参数)分别为0.691和12.626;在相同蓄积量条件下,林龄、单位面积蓄积量和面积也不同,因此b与总面积之乘积也不同;在相同蓄积量条件下,幼龄林单位面积蓄积量小,面积较大,总生物量较大;而成熟林、过熟林的单位面积蓄积量大,面积较小,总生物量较小,因此该模型在一定程度上体现了林分蓄积量和面积差异对估算结果的影响。计算结果表明马尾松总生物量为142.82 Tg,总碳储量为74.24 Tg。换算因子连续函数法是基于蓄积量和面积进行计算的方法,数据较为容易获取,通常无需进行大规模野外实地调查;该方法适用于多种尺度,具有一定普适性,可估算大部分森林类型的碳储量,但由于某些森林类型样本不足,且将生物量和蓄积量看作一种简单的线性关系,该方法尚存在较大争议。
-
马尾松木材密度SVD为0.448(t.d.m·m-3),综合BEF为1.294,采用根茎比0.173,计算得到总生物量。计算结果表明重庆市马尾松总生物量为116.65 Tg,总碳储量为61.49 Tg。其本质是基于调查数据进行分析后再扩展到整个森林区域,同时采用能够准确反映蓄积量与林分年龄关系的生物量扩展因子,更贴合森林实际情况[30-31]。但与上述两种方法相比,扩展因子法计算过程更为复杂,且对数据精度要求较高;若森林类型有误、数据缺失或参数设置不精确,则易产生较大误差。
-
VEFM中马尾松具体参数详见表 2,将BEF综合值与R替换为分龄组的参数,SVD与BEFM一致(0.448 t.d.m·m-3),总生物量即为各树种(组)各龄组生物量的总和。BEF值变化反映森林生长发育的不同阶段,定期监测其变化可发现病虫害、自然灾害等潜在问题,并采取相应干预措施[30, 32]。BEF在马尾松中随林龄增加逐渐减小,从幼龄林1.381降低到过熟林1.285。R值是指根系生物量与地上生物量之比,反映植物根系与地上生物量的分配关系:高R值代表根系较为发达,能更好地吸收水分和养分,其抗逆性及稳定性较强[32]。不同龄组之间R值变化较大,最高达0.231,近熟林阶段R值最高。
一般来说,幼中龄马尾松正处快速生长阶段,光合作用效率较高,此时植被对CO2吸收固定能力强,有机物质积累较多。幼龄林阶段生物量(23.78 Tg)与碳储量(12.53 Tg)增长迅速,但总体水平仍较低;中龄林阶段马尾松生物量(64.82 Tg)和碳储量(34.17 Tg)逐渐增加;近熟林阶段达到较高水平,生物量为29.44 Tg,碳储量为15.52 Tg;成熟林阶段因重庆市成熟马尾松林较少,其生物量仅为6.40 Tg,碳储量为3.37 Tg;过熟林马尾松碳储量(0.10 Tg)和生物量(0.18 Tg)均低于其他龄级,这主要是由于生长速度减缓、林分结构变化、自然和人为干扰以及森林演替等因素共同作用的结果。过熟林阶段生长速度显著减缓,生物量积累相应减少,光合作用效率降低,年净固碳量相对较低,同时过熟林处于森林演替后期阶段,森林结构逐渐老化或被其他树种替代,更易受自然干扰(如风倒、病虫害等导致树木死亡)和人为干扰(如采伐、森林火灾等破坏森林结构)。不同龄组马尾松总生物量为124.62 Tg,总碳储量为65.69 Tg,表明分龄组的VEFM更符合林分生长规律。
-
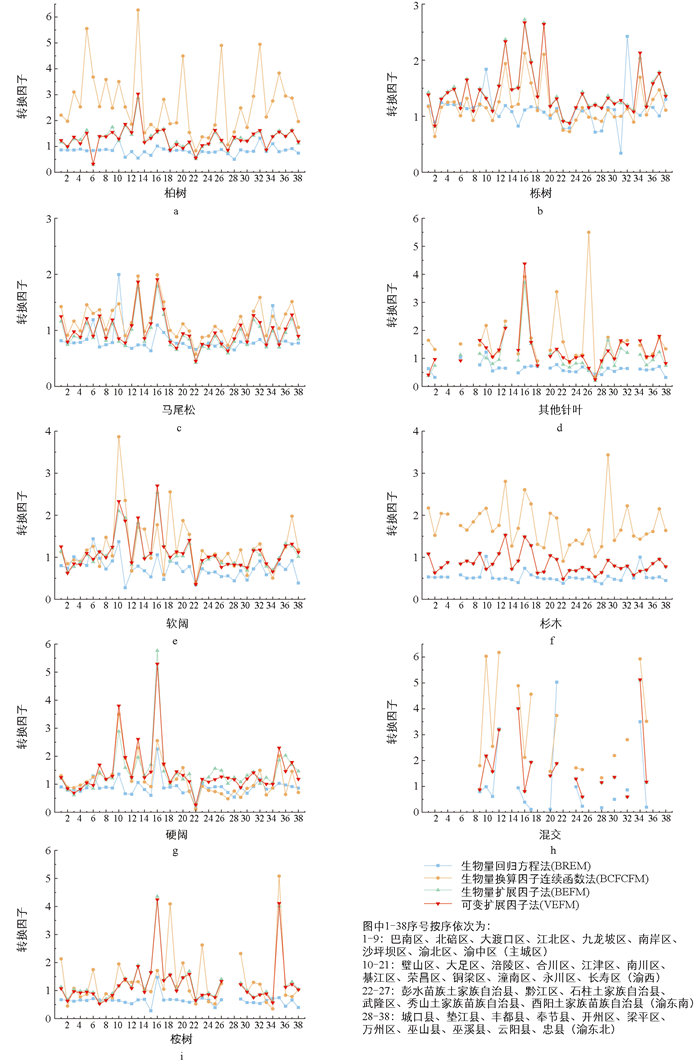
4种模型计算方法的稳定性可以从各树种的转换因子f值角度分析(图 3),f为生物量与蓄积量之比,反映单位蓄积量对应的生物量大小。总体来看,f值在不同方法下由高到低依次为换算因子连续函数法(1.19)、可变扩展因子法(1.05)、扩展因子法(0.98)、回归方程法(0.79)。整体上看,全部参考实测数据的BREM各林分类型的转换因子均小于其他3种模型,各片区内部相对变化幅度小;BCFCFM由于与单位面积蓄积有关,继而与采伐等生产经营活动有关,特别是在树种调节参数较大时,造成林分类型在不同实验区域转换因子波动最大,即稳定性较差;BEFM与VEFM的转换因子在数值上具有高度相似性,除个别区县外表现出良好的稳定性。例如生物总量最大的渝东北区,4个模型所得生物量(对应转换因子f值)依次为84.45 Tg(0.82)、139.03 Tg(1.35)、115.30 Tg(1.12)、120.21 Tg(1.17)。而使用同方法对同实验区域同树种计算,即使是胸径相近的2株柏树其总生物量和转换因子亦存在较大差别,这说明不同个体间生物量变动范围较大。因此,仅用一株平均标准木生物量衡量估测整个小区域生物量,可能带来较大误差。主要源于树种生长特性与生长环境有别,且受立地条件(坡度、地形等)、生产经营方式以及是否纯林等因素影响,生长速度较快、生物量积累较多的纯林其BEF值相对较高。针叶林虽然BEF值不一定高于阔叶林,但其火灾风险相对较高。这是由于针叶林木材密度较大,单位蓄积对应的生物量较多,火灾时枯枝落叶等可燃物载量大,加之树冠结构密集,使得火势迅速蔓延难以控制,防火难度高。因此,针对火灾风险较高的针叶林,可增加防火隔离带设置,定期清理林下可燃物,减少燃料积累,同时采取适当的森林经营措施。此外,BEF随林龄增大而降低,幼龄林BEF值较大,火灾风险较高,需进一步加强防火措施。
-
将估算出的乔木林生物量、碳储量及碳密度数据与其他学者使用不同模型获取的结果进行对比(表 5)。王义贵等[33]通过实测树高与胸径数据构建树木异速生长方程,计算林木地上生物量(干、枝、叶生物量之和),并进一步换算得到林木总碳储量。廖明蓉等[34]利用两次森林资源二类清查数据与林业工程数据,运用换算因子连续函数法,在林分小班尺度上测定森林含碳量,估算样区2002-2012年间恢复计划实现的碳增汇贡献。李奇等[35]基于森林资源清查数据,针对各优势树种拟合单位面积蓄积与林龄的Logistic回归生长方程,采用IPCC材积源-生物量法估算我国该阶段乔木林碳储量。陈雅如等[36]通过基于本地参数化的Biome-BGC模型,以长期监测气象数据驱动模型,模拟三峡库区针叶林、针阔混交林、阔叶林与灌木林植被层、凋落物层、土壤层的净初级生产力(NPP)与碳密度。
结果对比表明,基于实测数据构建的VEFM方法更适用于重庆市全市及县域尺度下乔木林碳储量和碳密度的计算。在不考虑其他外界因素影响的情况下,按平均碳密度年增长率1.08%估算,其他学者各阶段各森林类型平均碳密度介于28.15~34.01 Mg·hm-2之间。对比发现,BCFCFM法计算的各类碳密度普遍高于上述范围,BREM法结果偏低。一方面可能是由于针叶林中包括马尾松林、柏木林、杉木林、温性松林以及针叶混交林,在模拟中均统一按马尾松林处理,造成数值偏高;另一方面,随着各乔木龄组结构发生显著变化,不同龄组的作用机制存在差异。
采用F统计检验对4种模型间精确度差异进行两两比较(表 6),其他学者报道的碳密度介于28.15~34.01 Mg·hm-2之间,与本研究结果较为接近。结果表明:BREM与BCFCFM、BREM与VEFM之间的拟合效果差异具有统计学意义,p值分别为0.002和0.044;其余方法两两比较差异均无统计学意义(p>0.05)。相较于BEFM,VEFM考虑不同林龄生物量差异,并引入可变BEF与R值进行校正,估算结果更具合理性。
2.1. 相对误差
2.2. 计算过程及模型优缺点
2.2.1. 生物量回归方程法(BREM)
2.2.2. 生物量换算因子连续函数法(BCFCFM)
2.2.3. 生物量扩展因子法(BEFM)
2.2.4. 可变扩展因子法
2.3. 稳定性
2.4. 精确度
-
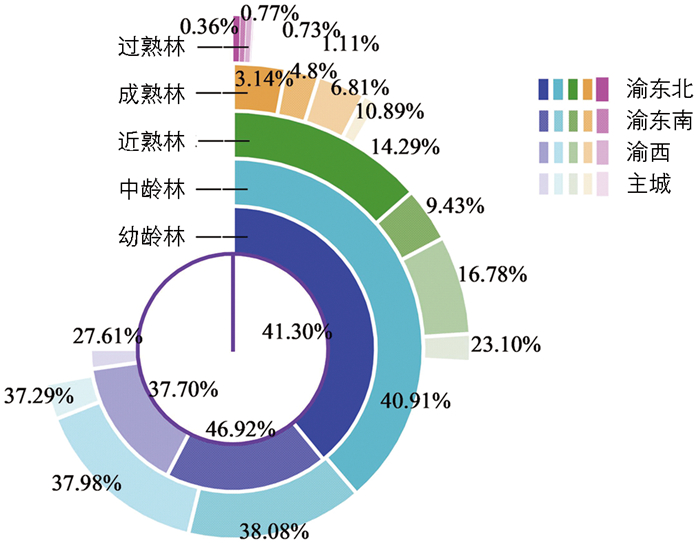
基于前述模型比较结果,后续分析主要采用可变扩展因子法的计算结果(表 7)。4大片区龄组构成(图 4)能够更好地反映出森林生长发育状况以及可采伐经营阶段。
-
林龄结构被认为是森林结构与功能的主要驱动因子,林龄与乔木林生物量、碳储量及碳密度紧密相关[37-39]。龄级的变化在很大程度上会影响森林碳动态,有研究表明森林生态系统碳储量随林龄的增长而增长,过熟林拥有较多的碳蓄积量,而幼龄林拥有较高的碳吸收率和较少的碳蓄积量[12, 40]。由表 7可见,基于可变扩展因子法计算,重庆市乔木林总碳储量为126.37 Tg,平均碳密度为35.54 Mg·hm-2,且马尾松碳储量在乔木林总碳储量中占据优势地位(约占51.98%)。从乔木林龄级分布看,5个龄级的碳储量分布不平衡。重庆市乔木林碳储量大小按龄组由高到低依次为:中龄林(58.22 Tg)、幼龄林(33.41 Tg)、近熟林(24.85 Tg)、成熟林(8.59 Tg)、过熟林(1.30 Tg),其分别占碳储量总量的46.07%、26.44%、19.66%、6.80%、1.03%。重庆市乔木林林龄结构以幼龄林和中龄林为主,其面积占全市乔木林总面积的80.60%(图 4),碳储量占乔木林总碳储量的72.51%,而近熟林和成熟林面积占全市乔木林总面积的18.09%,碳储量却占乔木林总碳储量的26.46%,由此可见重庆市乔木林碳储量具有较大的增加潜力。不同林龄组乔木林碳密度也存在较大差异:由低到高依次为幼龄林(23.60 Mg·hm-2)、中龄林(42.94 Mg·hm-2)、成熟林(49.50 Mg·hm-2)、近熟林(52.83 Mg·hm-2)、过熟林(54.62 Mg·hm-2),总体表现出林龄越大碳密度越高的趋势。
在自然生态系统中,不同林分类型在组成、物种稳定性、生长过程及生产力方面各不相同,造成其碳储量与碳密度各异[41]。阔叶林在演替顶级阶段地上植被所占比例为18%~44%,但其碳密度(29.83 Mg·hm-2)并不比针叶林高(38.00 Mg·hm-2),这一差异可能与针叶林具有较长的生命周期、较高的生物量积累能力以及稳定的生态系统结构有关。同时,马尾松碳储量是各片区乔木林碳储量的主体,分别占渝东北、渝东南、渝西、主城区同期乔木林碳储量的49.67%、46.17%、57.24%、72.45%;各片区栎树、柏树的碳储量也处于较高水平;马尾松、栎树、杉木、柏树的碳密度分别为44.74 Mg·hm-2、34.03 Mg·hm-2、26.25 Mg·hm-2、24.94 Mg·hm-2,这与林分面积和林龄密切相关。相较于其他3个片区,主城区的成熟林占比较高,这主要是由于主城区地势平坦,是主要的农业和工业生产基地,交通区位优势明显[42],在追求经济发展的过程中,除高山山脉地区保留了自然生态系统外,其余地区基本受到人为干扰,致使主城区的成熟林较多,但其自然生态空间受人为活动影响较大。
-
重庆市乔木林总体以天然林为主,不同区域因其区域面积、气候条件、地理分布及林业政策、林地开发保护和社会发展状况不同,乔木林碳储量以及碳密度大小存在明显差异,其地理分布格局具有重庆市特有的山地森林特征,各片区均以幼龄林和中龄林为主。乔木林碳储量按片区划分由高到低依次为:渝东北(61.37 Tg)、渝西(35.96 Tg)、渝东南(24.41 Tg)、主城(4.63 Tg)。碳储量主要集中在渝东北和渝西,整体呈现区域差异明显的分布格局。渝东北碳储量占全市总碳储量的48.56%,乔木林面积较大,但其碳密度(35.72 Mg·hm-2)与全市水平基本相当。而渝西地区乔木林面积占全市总乔木林面积的18.78%,但其乔木林碳储量却占全市总碳储量的28.46%,且碳密度(53.87 Mg·hm-2)明显高于全市平均水平;主城区由于社会经济发展迅速、人口密集,承担都市核心区功能,因此乔木林面积较小、占比较低,其碳储量也较小(4.63 Tg);渝东南碳密度最小(24.08 Mg·hm-2)。就全市来看,乔木林面积是森林碳储量大小的重要影响因素,乔木林面积较大区域碳储量也较大;而碳密度大小与区域单位面积蓄积关系密切,单位面积蓄积越大,其碳密度也越大(表 8)。
由表 8可知,各区县乔木林碳储量、碳密度分布格局在地域上也呈现出一定不平衡性,整体差异较为明显。中高海拔和坡度较陡区域是重庆市森林植被主要分布区和生物量集中分布地带,如奉节县(16.32 Tg)、綦江区(9.82 Tg)、城口县(9.62 Tg)、巫溪县(9.48 Tg)、武隆区(6.67 Tg)均为碳储量高值区。具体表现如下:
1) 渝东北地区碳储量较高:重庆渝东北地区乔木林碳储量高于其他片区。其中天然林碳储量较高地区主要分布在一些山区或林场,这些地区通常有较为丰富的自然资源和良好的生态环境。
2) 奉节县、綦江区、城口县、巫溪县、涪陵区碳储量排在前5位:依次为16.32 Tg、9.82 Tg、9.62 Tg、9.48 Tg、7.44 Tg,5个区县合计占全市乔木林碳储量的41.69%。碳储量呈现出内部分布不均的特点。生态条件优越、保护措施得当的地区,如部分自然保护区或风景名胜区,森林碳储量相对较高;而一些人类活动频繁、生态环境受到破坏的地区,碳储量相对较低。
3) 不同区县乔木林占比:按经济区域分,三峡库区生态经济区19个区县森林面积约占全市森林面积的80%,其乔木林碳储量也相应较高;而城市经济发达的9个区(即中心城区)以及部分渝西经济走廊区县,其乔木林碳储量相对较低,由此可见森林面积是森林碳储量大小的重要影响因素。
3.1. 乔木林林分结构和龄组碳储量分布状况
3.2. 乔木林碳储量及碳密度地理分布格局
-
本研究针对乔木林碳储量估算中模型缺乏系统评估的问题,以典型山地城市重庆为对象,通过集成森林资源清查数据,系统比较了4种主流估算方法的性能差异并揭示碳储量分布特征。结果表明:
1) 从方法角度看,BREM精度受原始数据质量限制,适用范围较窄;BCFCFM虽具有数据获取便利性和普适性优势,但其高波动性(f=1.19)可能导致跨区域应用效果不稳定,且生物量与蓄积量的简单线性关系假设忽略了林龄、密度等非线性影响,尚存在争议;相比之下,BEFM基于蓄积量、木材密度、生物量扩展因子和根茎比实现区域尺度扩展,能较好表征蓄积量与林龄的关系,而VEFM在此基础上引入动态参数体系(分龄组BEF与R值校正)综合表现最优,更符合林分生长规律,误差控制在±5%以内,这一优势源于其对林分动态变化的适应性,与Du等[43]在亚热带森林的研究结论一致。但其对数据精度要求较高,且参数敏感性较强,可能限制其推广应用,森林类型误判或数据缺失也会导致显著误差。因此,为进一步提高估算精度,建议融合激光雷达(LiDAR)冠层参数(如RH100)进行协同校正,该方法能有效量化三维冠层结构特征,弥补传统地面调查在垂直维度信息缺失的不足,特别是在复杂地形区域可降低因林分垂直异质性带来的估算偏差[44-45]。
2) 从空间格局看,重庆市碳储量的区域差异明显,渝东北中高海拔区为碳汇核心区,不仅受地形(坡度、海拔)调控,还与优势树种马尾松(贡献率51.98%)的空间分布密切相关。渝东北碳储量最高(61.37 Tg),占全市总碳储量的48.56%;渝西地区碳密度最高(53.87 Mg·hm-2),明显高于全市水平。这种差异主要受3个因素驱动:林龄结构失衡(经济快速发展地区往往伴随森林资源低龄化)、城市化导致的森林破碎化以及树种组成差异(自然保护区与人工林树种组成不同)。这一发现与Wu等[46]关于“地形—林分协同驱动碳汇空间异质性”的结论一致,表明未来碳汇管理需综合考虑立地条件与树种配置。然而,当前重庆市森林仍面临林龄结构不均衡(幼龄林和中龄林占比较高)和人为干扰(如历史砍伐、火烧及不合理经营活动)等挑战。因此,在碳汇管理方面提出以下建议:①保碳:在武陵山等高碳密度区建立碳汇保护区,实施禁伐政策维持原生森林碳库、促进自然增碳,同时科学经营人工林、减少碳泄漏风险,实现碳汇潜力的长期效益;②增碳:通过优化林分结构,如对占主导地位的马尾松林进行混交林改造、引入高固碳树种造林等提升森林碳储量和碳汇能力;③扩碳:在主城周边实施近自然改造提升碳积累潜力,在保障城市用地平衡的前提下将低效林地、退化地转换为高碳密度的近自然林,提升可持续碳汇功能。
 DownLoad:
DownLoad: