


 下载:
下载:
-
环境因子在时空上具有高度异质性,植物在不同生境条件下因光照、温度、湿度等的差异而表现出不同的生长和生理响应[1].凤丹,又名铜陵牡丹、铜陵凤丹,属江南品种群,其根皮有镇痛、解热、抗过敏、消炎、免疫等药用,具有根粗、肉厚、粉足、木心细、亮星多、久贮不变质等特色.由于种子油用的开发和推广,凤丹(Paeonia ostii)因植株高大,耐湿热、花量大、结实多、生态适应性强、病虫害少等优点成为当前油用牡丹的主栽品种[2-3].但是,目前对其研究主要集中在引种驯化、品种选育等方面,缺少生境微气候条件下对其生长、生理及光合特性的研究,丰产栽培技术缺乏理论支持和方向指引.
植株的生长性状是其对栽植地环境适应性的直观反映,光合作用是植物响应环境因子的重要方面.光照强度、温度和湿度等因子的变化对光合作用器官的形成及其功能存在显著影响,进而影响植物的净光合效率.已有的研究表明,在适度遮荫条件下牡丹净光合速率提高,植株成花率高[4].重庆地区栽植的凤丹多处于林下和空旷地两种不同的生境条件,但栽植效果尚无定论.开展上述两种生境条件下凤丹生长、功能叶叶绿素质量分数及光合日变化比较研究,可以对探索重庆地区凤丹栽植的适宜生境和优化栽培提供依据.
全文HTML
-
试验地位于重庆市巫山县两坪乡花梨坪(N 31°7′31″,E 109°58′35″),海拔825~850 m,坡度小于5°,属亚热带季风性湿润气候,年平均气温15.3 ℃,年平均降水量1 040 mm.设置巫山脆李(树龄为11年,平均株高4.5 m,郁闭度0.6,种植密度为630株/hm2)下和空旷地两种生境.两块试验地相邻,土壤均为中性偏碱紫色土,土层厚度60 cm;凤丹均为同一时间栽植,树龄8年,管护条件一致.于2017年6月初分别于该两种生境中设置5个5 m×5 m样地,进行不同生境条件下凤丹生长情况和光合特性测定.
-
不同生境条件下每块样地中随机选择6株,测定其株高、冠幅、一级分枝数及挂果量,计算单位冠幅面积挂果量.
-
叶片光合色素质量分数测定采用丙酮-乙醇提取法(V丙酮:V乙醇=1:1).称取0.1 g叶置于小容量瓶中,加入10 mL丙酮乙醇混合液,黑暗条件下浸提24 h.浸提液采用Spectrumlab 22可见光分光光度计分别在波长470 nm,645 nm和663 nm处比色,计算出叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)的质量分数[5].
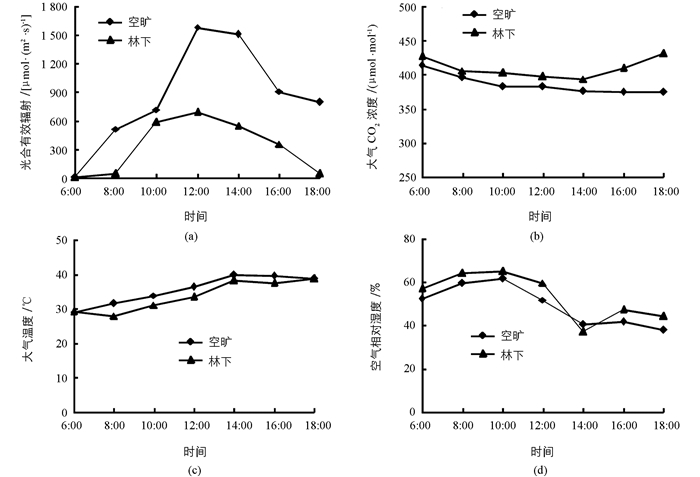
叶片光合参数日变化的测定采用Li-6400便携式光合作用测量系统(LI-COR,USA),在晴朗无风的天气自然条件下进行测定.两种生境条件下每块样地各选择2株,每株选取相同部分、生长健壮、无病斑的成熟叶测量.从6:00至18:00每2 h测定1次,每次测定记录6组数值,测定参数主要包括净光合速率(Pn),蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)及环境因子,包括光合有效辐射量(PAR)、大气CO2浓度、大气温度(Ta)、空气相对湿度(RH),试验期间的环境因子日变化见图 1.光能利用效率(LUE)为叶片吸收的每摩尔光合有效辐射总碳的固定量,即净光合速率与光合有效辐射(PAR)的比值.水分利用效率(WUE)为植物叶片每折腾一定量的水分所同化的CO2量,即净光合速率与蒸腾速率的比值.
-
试验数据用Microsoft Excel 2007软件进行录入、统计并作图,SPSS 19.0进行方差分析.
1.1. 试验方法
1.2. 生长指标测定
1.3. 光合作用相关指标测定
1.4. 数据统计与分析
-
如表 1所示,林下生境中的一级分枝数较空旷地高51.07%,差异具有统计学意义(p<0.05);前者冠幅面积和单位冠幅面积挂果量分别较后者高2.0%和21.57%,差异不具有统计学意义.空旷生境下树高较林下高4.40%,但差异不具有统计学意义.
-
如表 2所示,林下生境中凤丹植株功能叶的叶绿素a和叶绿素b质量分数较空旷生境下的差异具有统计学意义(p<0.05),前者在两项指标上分别高出后者103.53%和122.86%;而类胡萝卜素质量分数高40.0%,但差异不具有统计学意义.
-
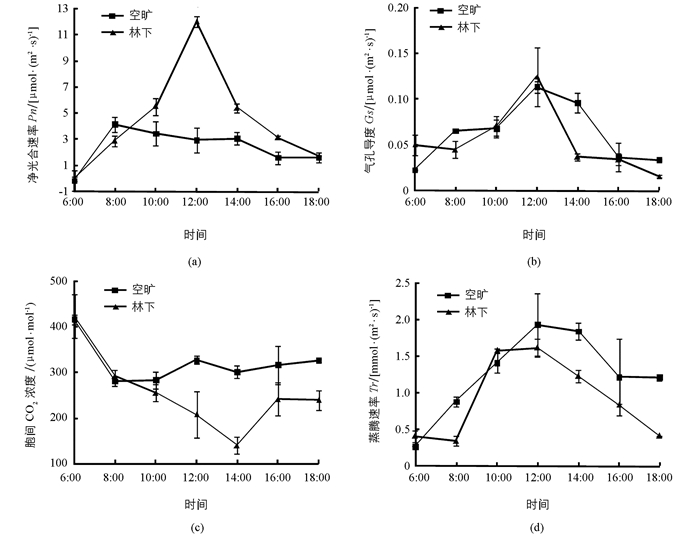
如图 2(a)所示,两种生境条件下的凤丹净光合速率(Pn)日变化趋势不同,林下生境中呈现明显的“单峰”曲线,在12:00时出现明显峰值,较其他时间段的差异具有统计学意义(p<0.05);而空旷生境下则呈现不明显的“双峰”,在上午8:00时出现第1个峰值后开始下降,下午14:00出现第2个峰值,比第1个峰值低25.78%,分析表明二者之间差异不具有统计学意义.就各时间段而言,林下生境的凤丹Pn普遍高于空旷生境,前者10:00至14:00之间的Pn显著高于后者(p<0.05). 图 2(b)和2(d)分别显示凤丹气孔导度(Gs)和蒸腾速率(Tr)的日变化特征.从趋势上看,林下生境的凤丹Gs日变化在12:00时出现峰值的“单峰”现象明显,而Tr在12:00时虽处于全天的最高点,但相较10:00时的差异不具有统计学意义;空旷生境下Gs和Tr变化趋势基本一致,均在12:00时出现全天最高值后逐渐下降,但峰值并不突出.各时间段内,空旷生境下凤丹的Gs和Tr总体高于林下生境,这可能与外界较强的光强和温度有关. 图 2(c)显示出林下生境凤丹叶片胞间CO2浓度(Ci)日变化均呈现明显的“单谷”曲线,在14:00时呈现最低值,总体上林下生境下的Ci低于空旷生境.
-
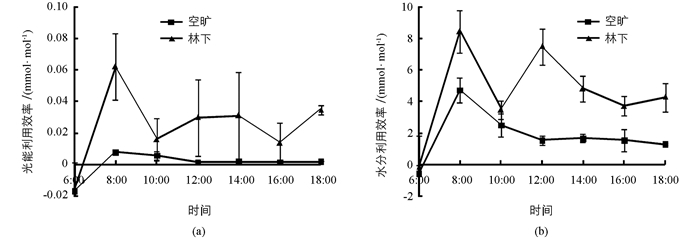
如图 3所示,两种生境条件下凤丹的光能利用效率和水分利用效率变化趋势均不同,但相同生境下二者的变化趋势基本一致.两种生境的凤丹光能利用效率和水分利用效率均在8:00时出现最高值,林下生境凤丹的全天变化趋势波动较大,而空旷生境的自最高值后呈平稳下降趋势.总体上看,各时段林下生境凤丹的光能利用效率和水分利用效率均高于空旷生境,除6:00和10:00外,林下生境与空旷生境差异具有统计学意义(p<0.05).
2.1. 不同生境条件下凤丹生长情况
2.2. 不同生境条件下凤丹叶的光合色素质量分数
2.3. 不同生境条件下凤丹光合日变化情况
2.4. 不同生境条件下凤丹光能利用效率和水分利用效率日变化情况
-
研究结果显示,相较长时间裸露在自然光照下,适当的林下生境(遮荫)条件能够降低夏季光强对凤丹的照射,减少光抑制现象发生.虽然这在一定程度上会使林下生境中的光合有效辐射减少,但凤丹长期的环境适应和自身的生理调节反而提高了其光能利用效率和净光合速率,保证了光合作用的正常进行.因此,合适的林下生境(适当遮阴)可能更有利于凤丹生长.
有研究指出,凤丹单株坐果量和单位冠幅面积产量较高的植株丰产性较好.田间选优标准以单株坐果量、单株籽粒数、籽粒千粒质量3个产量决定因素为主,宜选择树势旺盛、分枝多、无病虫害的单株建立种子园或母树林[6-7].本研究林下生境凤丹一级分枝数、单株挂果量、冠幅面积和单位冠幅面积挂果量均优于空旷生境,更具丰产性.
叶绿体色素是植物进行光合作用的物质基础,其质量分数高低直接决定了植物光合作用的强弱[8].野外试验中仅凭肉眼就能直观感受到林下生境下的凤丹叶片叶色浓绿,叶片偏大;而空旷生境下的叶片相对翠绿,叶片较小.试验结果显示林下生境凤丹功能叶叶绿素a和叶绿素b质量分数均显著高于空旷生境,而类胡萝卜素低于后者,这也验证了感官观察的结果.在光合作用中叶绿素a和叶绿素b主要起吸收光能的作用[9],虽然林下生境的光合有效辐射低于空旷生境,但也促使植株本身合成更多的叶绿体色素应对这一相对环境劣势.同时林下生境因遮荫,在一定程度上也缓解了水分亏缺和强烈太阳辐射因子胁迫的副作用,这也有利于叶绿体色素合成.
光合作用是植物生长发育的基础,为植物生长提供所需的物质和能量,而光合日变化直接反应了植物光合日生产能力,没有“午休”现象更能较大地提高植物光合日同化量[10-13].在本研究中,林下生境下凤丹净光合速率日变化在12:00时呈现明显的单峰曲线,而空旷生境下在这段时间可能存在轻微的“午休”.引起净光合速率变化的原因主要分为气孔因素和非气孔因素,前者是因气孔导度下降阻止了CO2的供应,只有气孔导度和胞间CO2浓度以相同的方式变化时,才能确定光合速率变化是由气孔因素引起的;而后者是因叶肉细胞光合能力下降使得CO2同化能力减弱,从而使胞间CO2浓度升高[14-15].本研究的空旷生境下正午时分气孔导度和胞间CO2浓度虽然变化趋势一致,但前者和蒸腾速率也是一致的,说明此时气孔开放程度较高,并不完全满足气孔关闭导致净光合速率下降的“午休”特点,可能还存在非气孔因素的原因.高温、强光照及干旱使得植株呼吸作用过强,消耗物质较多,导致细胞原生质结构破坏[16]可能是其中一个原因,但仍待进一步研究确定.林下生境下凤丹的净光合速率、光能利用效率和水分利用效率总体均高于空旷生境,这可能是遮荫条件下植物自身的生理适应抑或是其本身的特性使得光能和水分被更好地利用,光合效率较高,有机物合成多,分枝多,植物长势好,这与在茶花幼苗、西番茄中的研究结果一致[17-18],同时验证了林下生境下凤丹生长相关性状优于空旷生境.此外,本研究仅验证了林下生境(适当遮荫)对于凤丹栽植有利,但何种遮光程度最为合适及需光特性还需进一步研究和探索.