


 下载:
下载:
-
在我国,小麦是仅次于水稻的粮食作物,对保障粮食安全具有十分重要的作用.由于小麦育种过程中过分集中地使用一些具有优良性状的亲本,使得小麦育成品种的遗传基础日益狭窄,遗传脆弱性增强,对各种不良环境的抵御能力日益下降[1].小麦的近缘属是一个巨大的、弥足珍贵的基因资源库,蕴藏着非常丰富的遗传变异,具有许多栽培小麦所不具备的优良特性,如抗病、抗逆、耐贫瘠、高营养、高蛋白和大穗多花多粒等[2],从小麦近缘属中挖掘有益基因可以拓宽小麦遗传基础,丰富遗传多样性,因而利用近缘物种育种成为小麦育种的热点方向之一.黑麦属(Secale L.)是小麦的三级基因源,黑麦具有高产、广适、抗病性好、抗逆性强、籽粒和茎叶产量高、营养成分丰富等优点[3],蕴藏着改良栽培小麦抗性、产量、品质、环境适应性等性状的丰富基因资源,对于丰富小麦的遗传变异、拓展小麦的种质资源具有重要作用.小麦与黑麦远缘杂交获得的小黑麦是第一个人工创造的新物种,也是黑麦染色体导入小麦形成的典型双二倍体,由于小黑麦同时具有小麦和黑麦不同属的染色体,因而是研究异属染色体配对行为及相应遗传性状的理想材料[4].世界上第一个应用在生产上的八倍体小黑麦品种于1973年在贵州试种成功,具有穗大、小穗数多、生长旺盛、蛋白质含量高、广泛的适应性和抗逆性等优点,对解决贵州粮食问题做出了特殊贡献[5].李方安等[6]用Kustro黑麦与绵阳11小麦杂交,获得了小麦-黑麦双二倍体材料MK-25,再用绵阳11等小麦和MK-25回交,从回交后代中选育出6R单体附加系、6RL端体附加系及来源于6R单体附加系的小片段渐渗系,且发现这些附加系和小片段渐渗系材料都表现出了对白粉病免疫的特性,表明Kustro黑麦上的白粉病抗性基因渗入到了小麦遗传背景中,因而利用异源单体附加系等诱导外源有利基因的渐渗是可行的.
FISH技术是以荧光素标记的核酸片段探针与染色体上的特异序列进行杂交,通过荧光检测系统检测杂交信号从而确定探针序列在染色体上的杂交位点的技术.目前,FISH技术在染色体定位、基因克隆、遗传标记以及染色体畸变等研究中得到了广泛的应用.罗巧玲等[7]利用GISH和多色荧光原位杂交(mc-FISH)技术,对8份农艺性状优良的小麦-黑麦材料进行染色体组成分析,鉴定出3份为六倍体小黑麦(AABBRR),2份为八倍体小黑麦(AABBDDRR),1份为1RS·1BL易位系,其余2份不具有可见的黑麦染色体或染色体片段;同时发现其中的3份六倍体小黑麦与2份八倍体小黑麦所含的黑麦染色体不完全相同.李俊等[8]利用四川地方品种蓬安白麦子与秦岭黑麦杂交得到八倍体小黑麦CD-13(AABBDDRR),并通过顺序FISH-GISH技术,发现该八倍体小黑麦1RS端部与7DS端部发生相互易位,鉴定其携带1RS-7DS·7DL小麦-黑麦小片段易位染色体. Tang等[9]使用Oligo-pSc119.2,Oligo-pAs1和Oligo-pTa535等高度重复序列探针对中国春和绵阳11小麦进行双色FISH试验,创立了中国春和绵阳11小麦的A,B和D基因组染色体FISH核型模式图;并通过对八倍体小黑麦的顺序FISH-GISH试验,识别了每一条染色体.杨武娟[10]经同步的GISH和FISH技术鉴定小麦-黑麦衍生系N9436B,发现N9436B包含的两条黑麦染色体是一对1R染色体,为小麦-黑麦1R附加系;并采用Oligo-pSc119.2探针初步建立了奥地利黑麦的FISH核型.何道文等[11]根据黑麦特异重复序列pSc20H设计特异引物,用PCR方法从中国春小麦与秦岭黑麦杂交后代(BC2F4)共75份材料中筛选出30份含有黑麦成分的材料.本试验拟对小黑麦染色体进行顺序GISH-FISH等分析,明确黑麦(RR)染色体在小麦遗传背景中的存在形式,分析R染色体组的FISH特点,建立其FISH核型,分析小黑麦的染色体组成,为其在小麦遗传育种中的应用提供参考.同时,本试验有利于拓宽小麦遗传基础及创建新种质,为外源染色体的有效转移提供借鉴,加速优良小麦新品种的选育进程.
全文HTML
-
本试验植物材料为小黑麦、黑麦(RR)和野生二粒小麦(AABB);该小黑麦对白粉病、条锈病和叶锈病等小麦主要病害表现出高抗或免疫,同时又具有丰产的优点;这些纯系材料保存于国家小麦改良中心贵州分中心实验室.
-
将种子放入盛有蒸馏水的培养皿中浸泡24 h,待种子发芽露白后,倒掉蒸馏水,将其放在铺有湿润滤纸的培养皿中于27℃恒温箱培养,待根长至2 cm左右时,取根尖放入湿润打孔的离心管中用N2O处理2 h,用90%冰醋酸固定10 min,加蒸馏水润洗后切取根尖,水浴酶解滴片,用OLMPUS BX60显微镜镜检,选择分裂相好、染色体清晰的玻片进行后续试验.
-
本试验中黑麦和野生二粒小麦的DNA提取参照黄娟[12]方法,荧光原位杂交技术参考Han等[13]方法,Oligo-pAs1-1,Oligo-pSc119.2-1,pSc200和pSc250探针由Thermo Fisher Scientific(上海)公司合成.试验中的pSc200和pSc250探针可以识别小黑麦中的R组染色体[14],而Oligo-pSc119.2能够辨别R组的每一对染色体[15].镜检选择FISH效果好的细胞用Cellsens Standard摄像系统照相,选取5个以上分裂相好且染色清晰的细胞用于GISH和FISH分析,以得到准确可靠的数据.
1.1. 材料
1.2. 方法
1.2.1. 染色体制片
1.2.2. 荧光原位杂交分析
-
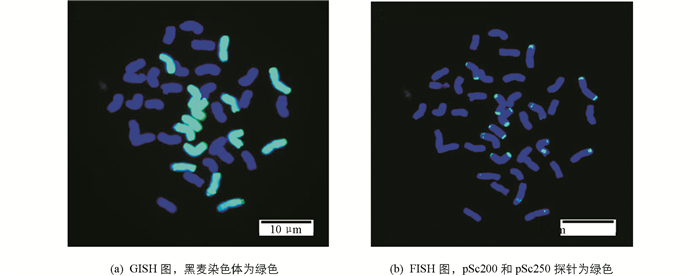
为了快速准确地区分小黑麦材料中的外源黑麦染色体,本试验分别以黑麦基因组DNA(绿)为GISH探针,以切断的野生二粒小麦基因组DNA为封阻,以pSc200(绿)和pSc250(绿)为FISH探针,进行了顺序GISH-FISH分析(图 1). FISH结果显示,小黑麦含有21对染色体,其中包括来源于黑麦的7对整条染色体,没有发现易位染色体;有2对染色体只在染色体长臂端部出现杂交信号,另外5对染色体的两端部都有杂交信号,但信号的强弱有所差异.该FISH和GISH结果一致,表明pSc200和pSc250为黑麦染色体端部特异重复序列探针,可用于小黑麦背景中黑麦染色体的快速检测.
-
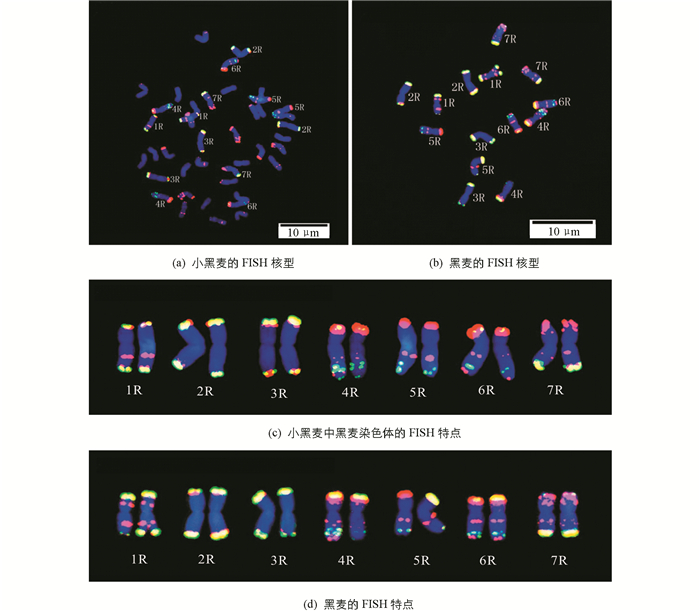
为了进一步识别小黑麦中黑麦的每一条染色体,以Oligo-pSc119.2-1(红),pSc200(绿)和pSc250(绿)为探针分别对小黑麦和黑麦材料进行了双色FISH分析,初步建立了小黑麦中R组染色体的FISH核型(图 2).结果显示,小黑麦的2R和3R染色体信号很相似,都只在端部有信号,但可依据着丝粒位置不同区分出2R和3R染色体(2R染色体着丝粒位于染色体亚中部,而3R染色体着丝粒在中部).通过对比小黑麦和黑麦的杂交信号,发现Oligo-pSc119.2-1探针在两个材料中的信号位点基本相似,只有小黑麦1R染色体短臂丢失了中间的信号. pSc200和pSc250探针在小黑麦和黑麦两个材料中的信号差异较大,小黑麦的信号相对黑麦有减弱或丢失现象,如1R染色体短臂近端部、4R染色体短臂端部、6R染色体两端部的信号都出现减弱;7R染色体短臂端部的信号丢失;而5R染色体长臂上的信号却增强了;并且pSc200和pSc250探针只在5R和7R长臂端部出现杂交信号.由上述可知,小黑麦中几个R组染色体的FISH信号发生了一些变化,其原因可能是在小黑麦形成过程中染色体结构发生了改变,从而使小黑麦的DNA重复序列呈现多态性.
-
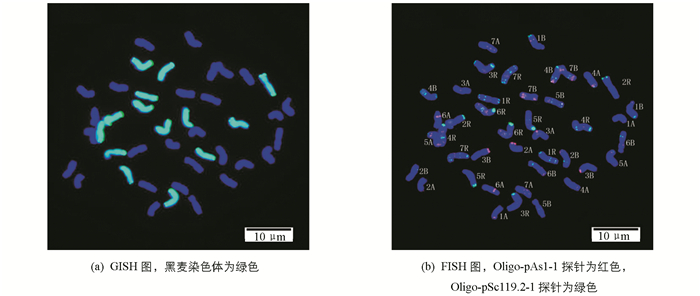
本试验以黑麦基因组DNA(绿)为GISH探针,以切断的野生二粒小麦基因组DNA为封阻,以Oligo-pAs1-1(红)和Oligo-pSc119.2-1(绿)为FISH探针进行顺序GISH-FISH分析(图 3),参照中国春(AABBDD)等小麦[9]和黑麦[10]的FISH核型,发现该小黑麦的染色体组成为AABBRR,即小黑麦的染色体构型为2n=42=28(AB组)+14(R组).该小黑麦的A组和B组染色体信号与已报道的硬粒小麦(AABB)[16]和中国春小麦[9]等都有所差异.与中国春小麦对比发现,该小黑麦的A组和B组染色体Oligo-pAs1-1的杂交信号整体增强,特别是5A短臂近端部、6A长臂中间和3B短臂端部出现明显的杂交信号;而Oligo-pSc119.2-1的信号出现减弱或丢失现象,例如,5A,2B和3B短臂端部都有信号丢失或减弱现象.与硬粒小麦相比,发现Oligo-pSc119.2-1杂交信号减弱或丢失明显,这可能是因为在小黑麦形成过程中,其染色体结构发生了变异.
2.1. 小黑麦中R染色体的鉴定
2.1.1. 小黑麦的顺序GISH-FISH分析
2.1.2. 小黑麦中R染色体的FISH核型
2.2. 小黑麦的染色体组成
-
通过远缘杂交将小麦亚族植物优异基因渗入到小麦遗传背景中,能够快速有效地检测和跟踪外源染色质.进行普通细胞学观察和GISH技术不仅能快速检测小麦背景中的外源遗传物质,还能揭示外源染色体在遗传上的不稳定对远缘杂交后代遗传稳定性的影响,在理论研究和育种实践中都有非常重要的意义[17].杂交技术根据所用探针不同,可分为GISH,FISH,物种专化重复序列原位杂交和单拷贝或寡拷贝序列原位杂交等[18].本试验利用GISH技术,快速区分小黑麦中的外源R组染色体. GISH技术能直观准确地检测出外源染色体及其片段,但无法快速有效地鉴定出具体的外源染色体,以及与这些外源染色体发生交换或重组的具体小麦染色体,尤其是难分辨亲缘关系比较近的染色体组,而重复序列探针的FISH技术就具有一定的互补优势.本试验利用双色FISH技术能够在染色体水平上准确辨别来源于黑麦R组的每一条染色体. pSc200和pSc250探针为黑麦端部特异重复序列,只在R组染色体端部出现杂交信号,这有利于快速高效地识别小黑麦中的R组染色体;而Oligo-pSc119.2-1杂交信号较强,其信号在小麦、黑麦染色体上都有分布,且能识别R组每一条染色体.
Alkhimova等[19]利用重复序列pSc200为探针,对一整套中国春-帝国黑麦二体附加系进行研究,发现黑麦染色体1R,2R,3R,5R和7R的端部或亚端部都存在变异.符书兰[20]分别利用Kustro黑麦、AR106BONE黑麦与绵阳11小麦杂交,获得的两种双二倍体材料进行FISH试验,发现在来自不同组合的小麦-黑麦双二倍体中,pSc200探针在黑麦染色体上的杂交信号不同;另外,其杂交信号的增减在所观察的细胞中都表现一致,证明探针pSc200的变异不是随机的,其结果还表明外源种质的供体种与受体种之间的确存在相互作用,且这种相互作用在多倍化早期就开始了.蒋敏[21]将绵阳26小麦与秦岭黑麦杂交得到F1种子,经人工加倍后获得双二倍体材料MQ,对MQ与绵阳26小麦的杂种F1代进行了分析,发现黑麦染色体6R长臂亚端部的pSc119.2信号出现了明显的缺失,黑麦基因组重复序列发生了变异,变异区域位于6R染色体长臂的亚端部.葛群等[22]利用GISH和FISH相结合的方法,对绵阳11小麦和抗病的威宁黑麦杂交、回交后代进行试验,分别鉴定出一个1R和5R单体附加系,发现黑麦染色体1R和5R单体附加系可以诱导小麦染色体变异.本试验小黑麦中1R,4R,5R,6R和7R染色体的FISH信号出现减弱、增强或缺失等变异情况,这与Alkhimova等[19]、符书兰[20]和蒋敏[21]的研究存在相似之处;其原因可能是在小黑麦多倍体化过程中造成基因组DNA含量的显著扩大,引起基因组的不稳定,导致DNA重复序列发生了变异,从而拓宽了小麦遗传背景,丰富了小麦种质资源.重复序列在植物基因组中占有很大比例,重复序列的扩增和分化导致了植物基因组进化[23].本试验小黑麦是由一种四倍体小麦(AABB)和黑麦(RR)经远缘杂交以及染色体加倍后获得的六倍体小黑麦(AABBRR)新种质,新形成的多倍体会通过序列的大量丢失,尤其是重复序列的大量丢失等方式控制DNA含量,从而更有利于新合成的多倍体在自然界的存活与进化.该种质种植于贵州高海拔生态区,粒大饱满,次生根系发达,顶二叶叶面积较大,根据王志伟等[24]对高海拔生态区不同小麦品种叶面积、叶绿素相对质量分数、根系性状及产量的研究结果,该种质具有较大增产潜力,这有待后续深入研究.