


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
植物生长受到环境因素的限制,其中气候是决定全球植被分布的重要因素之一[1]. 近年来,气候变化日益加剧. 联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)第六次评估报告指出,自第四次工业革命后,全球气候变暖的趋势更加明显,预计到2030年左右,全球地表温度相较于工业革命前将上升超过1.5 ℃[2]. 气候变暖影响全球水热循环,导致降水的时空分布不均,极端气候事件频繁发生[3]. 作为森林生态系统的基础,植物物种维持着森林生态系统的功能和稳定性[4-5]. 光照、温度、水分、土壤等环境因素的区域性变化将重塑植物群落[6-7]. 环境胁迫促使植物调整其生态特征和地理分布格局,逐渐扩散或迁移到更适宜的生境,以缓解不利环境条件的影响[8-10]. 因此,了解植物物种在大尺度范围内的分布模式及其在气候变化情景下的变化趋势,已成为生物地理学和全球变化研究的热点领域,对于生物多样性保护和森林经营管理具有重要的理论和实践意义.
物种分布模型(Species Distribution Models,SDMs)可确定单个物种或群落的环境生态位和潜在分布,目前被广泛用于预测气候变化下物种的地理分布模式[11-12]. SDMs的本质是通过数学统计方法建立物种发生数据与相应空间特征信息之间的关系,以分析物种对环境的耐受差异[13]. 基于不同理论和算法,常见的模型有Bioclim(生物气候分析系统)[14]、ENFA(生态位因子分析模型)[15]、GARP(遗传算法模型)[16]、GLMs(广义线性模型)[17]等物种分布模型. 然而,环境因子之间的多重共线性以及物种间的相互作用会降低部分模型的性能,导致预测精度降低或过度拟合的问题[18]. MaxEnt模型基于最大熵原理,以目标发生记录和环境特征值作为约束条件,通过最大熵概率分布判断目标概率分布[19]. MaxEnt模型对缺失的物种发生记录不敏感,具有高预测精度和较小的失真度,在样本量较小的情况下即可获得良好的预测结果[20-21]. 近年来,MaxEnt模型广泛应用于植物适生区变化的预测,如红松(Pinus koraiensis)、马尾松(Pinus massoniana)和沙棘(Hippophae rhamnoides)等物种.
柃属(Eurya)植物隶属于山茶科,为小乔木或常绿灌木[22],以树形优美而著称,适合作为观花和观果园林植物,可栽培于园林花坛. 柃属植物种类繁多,分布面积广,全球范围内有130种,我国境内就有80多个物种,是长江以南亚热带森林中的常绿灌木[23]. 柃属植物的枝叶可药用,具有清热、消肿的功效,同时也是重要的冬季蜜源. 柃属植物花粉充足,产蜜量丰富,对于蜂群安全越冬和春季的快速繁殖具有重要作用[24]. 目前,对于柃属植物的研究集中在种质资源调查[25-26]、营养繁殖栽培[27-28]等方面. 然而,在未来全球气候变暖背景下,环境压力可能会驱使柃属植物逐渐迁移,从而改变其地理分布格局. 岗柃(Eurya groffii)、柃木(Eurya japonica)和细枝柃(Eurya loquaiana)是中国常见的3种柃属植物,本研究以这3种植物为研究对象,利用MaxEnt模型预测其在当前及未来不同气候情景下的潜在分布区域的动态变化,并绘制柃属植物在中国境内分布的高分辨率地图,探究影响其分布格局的关键驱动因素,以期为柃属植物的资源保护和管理开发提供科学依据.
全文HTML
-
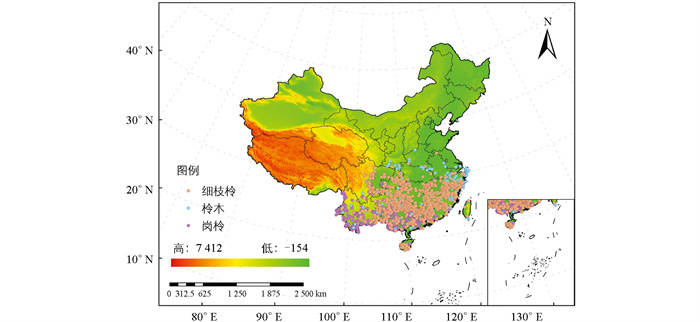
本研究以柃属3个物种(岗柃、柃木和细枝柃)为研究对象,并获取这些物种的地理分布点数据,数据来源包括中国数字植物标本馆(
https://www.cvh.ac.cn/ )、教学标本资源共享平台(http://mnh.scu.edu.cn/ )、全球生物多样性信息网络(http://www.gbif.org ). 对于具有地名但缺乏坐标信息的样点,通过百度地图获取经纬度坐标,并排除错误和重复的样点. 为避免样点间的空间自相关导致模型过度拟合,使用ArcGIS中的SDMToolbox v2.5工具包对样点数据进行稀疏化处理[29]. 具体而言,在每个地图栅格(5 km×5 km)内最多保留1个样点,最终确定了岗柃的535个分布点、柃木235个分布点和细枝柃741个分布点(图 1). -
环境数据共选取28个环境变量作为预测变量,包括19个气候因子、6个土壤因子以及3个地形因子(表 1). 当前气候因子(1970-2000年)的数据来源于世界气候数据库(
https://www.worldclim.org/ ),分辨率为2.5 min. 基于第六次耦合模式比较计划(CMIP6),未来气候因子(2080-2100年)使用了北京气候中心开发的BCC-CSM2-MR气候系统模式中3种共享社会经济路径(SSPs)数据,分别为SSP245(低等胁迫情景)、SSP370(中等胁迫情景)和SSP585(高等胁迫情景). 这3种情景分别假定2100年辐射强迫稳定在约4.5 W/m2,7.0 W/m2,8.5 W/m2[30-31]. 研究表明,BCC-CSM2-MR模式在对东亚夏季风和降水的模拟方面表现良好,并且对中国地区的气候预测具有较高的精度[32-33]. 地形因子数据来源于地理空间数据云(http://www.gscloud.cn ). 土壤因子数据来源于世界土壤信息数据库(https://soilgrids.org/ ),缺失的土壤理化性质数据通过计算邻近栅格的平均值进行补全[34]. 最后,使用ArcGIS v10.8对所有环境变量的栅格底图进行重采样,以确保具有相同的空间分辨率(5 km×5 km).由于环境变量之间存在强相关和多重共线性等问题,为避免过度拟合并提高预测精度,对环境因子进行筛选和剔除,以减少冗余信息对模拟结果的影响. 首先,在R4.2.1中,使用car数据包中的“vif”函数检验环境因子之间的多重共线性,设置方差膨胀因子(Variance Inflation Factor,VIF)阈值VIF<10. 使用“cor”函数进行了Pearson相关性分析,对于相关系数r>0.8的变量,结合植物的生存环境需求和生理特性,优先保留贡献率更大的变量. 岗柃和柃木各保留8个环境变量,细枝柃保留9个环境变量(表 2).
-
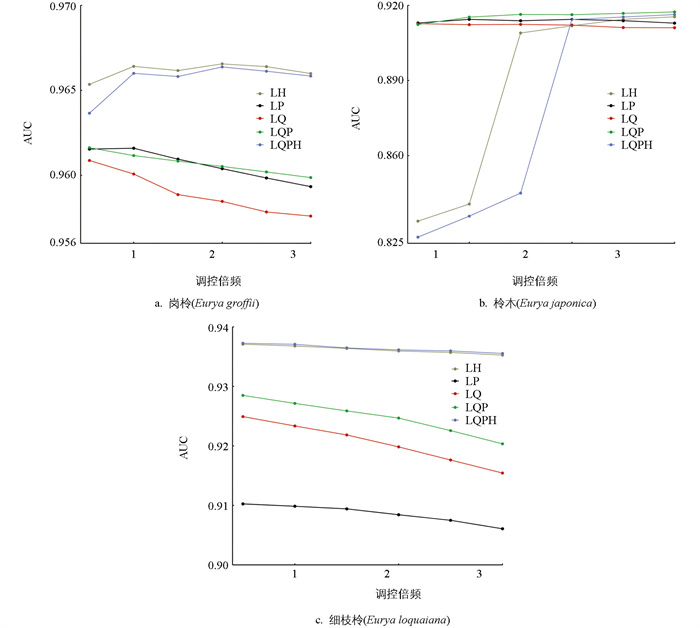
为了使岗柃、柃木及细枝柃的出现概率接近正态分布,选择75%的记录点数据作为训练集,而剩下的25%记录点作为测试集[35]. 利用R4.2.1中的ENMeval数据包对MaxEnt模型进行优化. 在模型优化过程中,修改和调试模型的特征和调控倍率2种参数[36]. MaxEnt模型提供了5种特征,包括线性(Linear,L)、二次(Quadratic,Q)、片段化(Hinge,H)、乘积型(Product,P)和阈值(Threshold,T). 本研究共测试了5种特征组合(Feature Combination,FC),分别为LP,LQ,LH,LQP和LQPH. 调控倍频设置为0.5~3,间隔为0.5,总共测试6种调控倍频[37]. 将上述参数进行组合测试,采用受试者工作特征(Receiver Operating Characteristic,ROC)曲线分析法对预测结果进行精度评估,以ROC的曲线下面积(Area Under the Curve,AUC)作为模型精度的评价指标.
参数优化完成后,选择具有最高AUC得分的参数组会用于最终的MaxEnt模型构建. AUC的取值范围为0~1,得分越高表示模型的可行度越高,预测效果越好. 根据AUC的划分,当AUC<0.6时,表示预测失败;当0.6≤AUC<0.8时,表示预测良好;当0.8≤AUC<0.9时,表示预测结果优秀;而当AUC>0.9时,表示预测非常准确[38].
-
本研究使用MaxEnt模型模拟产生的结果,通过ArcGIS v10.8转换为栅格数据并进行可视化处理. 利用重分类(Reclassify)工具,采用自然间断分级法(Jenks' Natural Breaks)将潜在适生区划分为4个等级:非适生区(<0.2)、低适生区(0.2~0.4)、中适生区(0.4~0.6)和高适生区(0.6~1.0). 同时,确定不同适生区占比,并计算了3种柃属植物在不同适生区的面积[39].
-
为分析物种适生区的空间格局变化,本研究将逻辑值L≥0.2的部分定义为潜在适生区,并赋值为“1”;将L<0.2的部分定义为非适生区,并赋值为“0”. 然后,在ArcGIS v10.8中将图层转换为二进制图层. 本研究将物种适生区的变化分为4种类型:非适生区、收缩区、保留区和扩张区. 3种柃属植物未来的分布面积变化均基于当前气候情景下该物种的潜在适生区面积来进行比较计算. 将未来适生区的二进制图层数值乘以2,并与当前适生区的二进制图层相减. 其中,0→0代表非适生区,1→0代表收缩区,1→1代表保留区,0→1代表扩张区. 通过这一可视化方法,能够实现3个柃属物种潜在适生区空间格局变化的可视化[40].
1.1. 物种地理分布数据来源
1.2. 环境变量的收集与筛选处理
1.3. 模型的构建优化及评估
1.4. 物种潜在适生区等级划分
1.5. 物种适生区空间格局变化
-
3种柃属植物的最优模型参数各不相同(图 2). 当FC为LH,调控倍频为2时,岗柃的AUC值达到最大,为0.967;当FC为LQP,调控倍频为2时,柃木的AUC值为0.917;细枝柃在FC为LQPH,调控倍频为0.5时,AUC值为0.937. 模型精度评估结果显示,3种柃属植物的AUC值均大于0.9,说明预测结果具有很高的准确性. 由此可见,模型具有较高的可靠性,能够很好地模拟气候变化条件下中国3种柃属植物的潜在适生区.
-
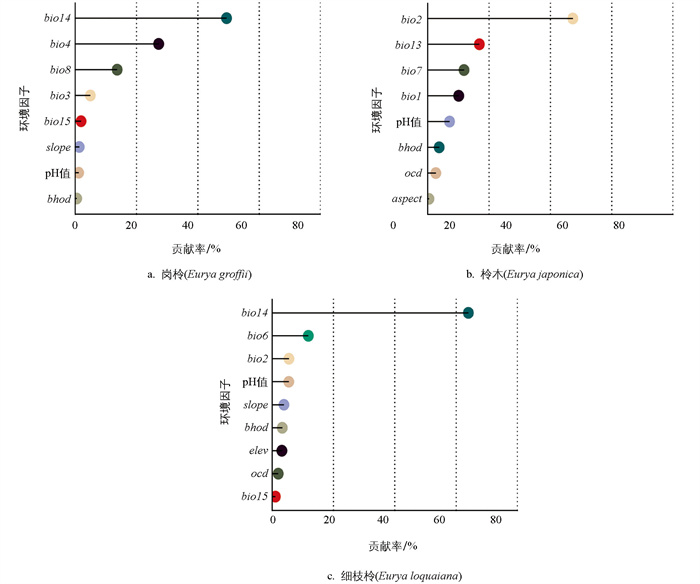
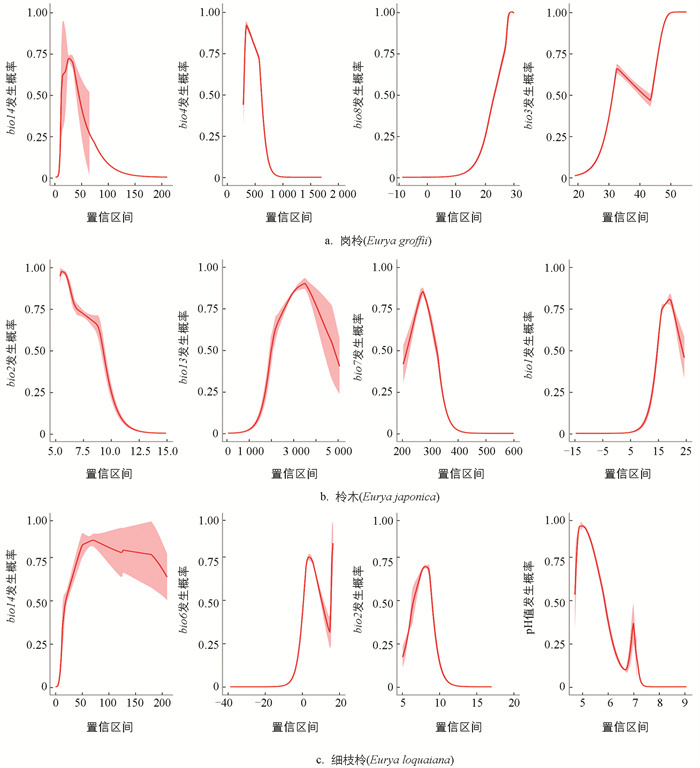
3种柃属植物的潜在分布由环境因子驱动,不同物种对主导环境因子的响应存在差异. 岗柃最主要的4个环境因子的贡献率累计达95%(图 3a). 其中,最干月降水量(bio14)的贡献最大,占比49.4%;气温季节性变化系数(bio4)占比27.2%;最湿季度平均温度(bio8)占比13.7%;等温性(bio3)占比5%. 响应曲线结果表明,岗柃的适生概率对最干月降水量和气温季节变化系数呈单峰模式. 最干月降水量的适宜区间为0~150 mm,最适值约为30 mm;气温季节性变化系数的适宜区间为0~500,最适值约为400. 此外,岗柃的适生概率随着最湿季度平均温度与等温性的增加而逐渐增大(图 4a).
影响柃木潜在分布的关键驱动因子的贡献率累计达86.2%(图 3b). 其中,平均气温日较差(bio2)贡献最大,占比47.3%;最湿月降水量(bio13)占比16.9%,适宜区间为100~500 mm,最适值约为350 mm;气温年较差(bio7)占比11.9%,适宜区间为200~400 ℃,最适值约为300 ℃. 年平均气温(bio1)的贡献占比10.1%,适宜区间为10~25 ℃,最适值约为20 ℃. 其中,柃木的适生概率随着平均气温日较差的增加呈单调递减,最适值约为5 ℃(图 4b).
影响细枝柃潜在分布的关键驱动因子的贡献率累计达86.7%(图 3c). 其中,最干月降水量(bio14)对细枝柃的分布影响最大,贡献率为64.0%;最冷月最低温度(bio6)为11.9%;平均气温日较差(bio2)为5.5%;土壤酸碱度(pH值)为5.4%. 细枝柃的适生概率随着最干月降水量的增加而升高,并在约75 mm时达到最大值;最冷月最低温度的适宜区间为0~20 ℃,平均气温日较差的适宜区间为5~10 ℃,土壤pH值的适宜区间为5~7,并在pH值约为5.2时适生概率达到最大值(图 4c). 综上所述,气候因素对中国3种柃属植物的潜在分布比土壤和地形因素具有更大的决定作用.
-
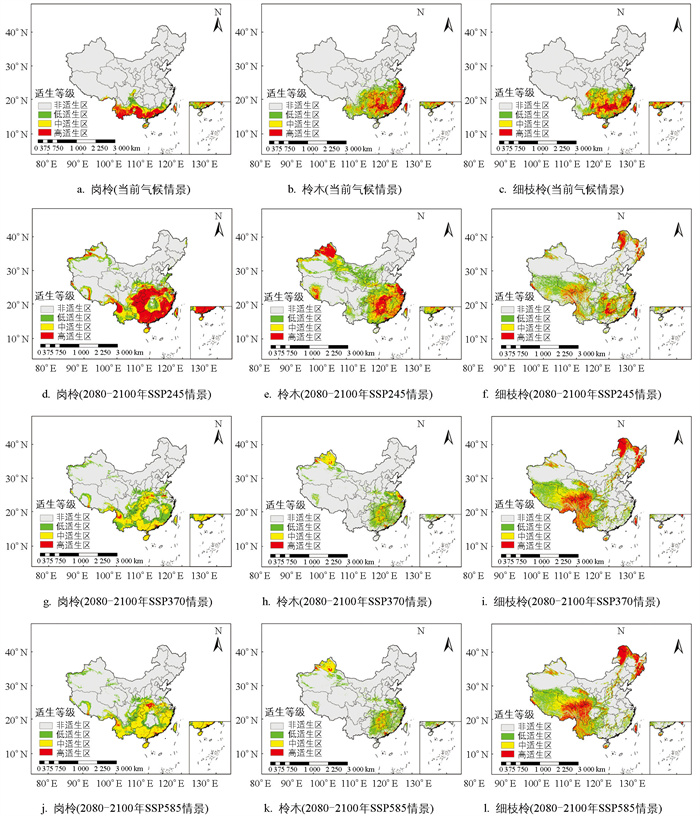
通过MaxEnt模型模拟,分别绘制了岗柃、柃木和细枝柃在当前气候条件及2080-2100年SSP245,SSP370,SSP585情景下的高分辨率潜在适生区分布地图. 在当前气候情景下,岗柃的适生区分布在中国的南部地区,主要集中在云南、广西、广东、福建、海南、中国台湾等地,总面积为9.54×105 km2(表 3),其中高适生区占比48.4%,中适生区占比23.6%,低适生区占比27.9%. 在未来3种气候情景下,岗柃的适生区面积均呈扩大趋势,高适生区面积则呈现单峰模式,即先增加后减少. 在SSP245的气候情景下,岗柃的高适生区面积达到峰值,为1.47×106 km2. 在2080-2100年的潜在分布区,岗柃主要集中于中国的华东、华南、华中地区,少量分布于甘肃、西藏、新疆等地(图 5).
在当前气候条件下,柃木的潜在适生区主要分布于华东、华中和华南地区,西南地区也有分布(图 5),总面积为2.00×106 km2,其中高适生区占比30.4%,中适生区占比28.7%,低适生区占比40.8%(表 3). 在未来气候情景下,柃木的适生区总面积和高适生区均呈先增加后减少趋势. 在2080-2100年SSP245情景下,适生区面积增加,而在SSP370和SSP585情景下,总面积减少. 在2080-2100年SSP245情景下,高适生区面积最大,为1.01×106 km2,主要集中于湖北、湖南和东部沿海地区. 此外,西北部地区包括甘肃、新疆、西藏等地也见分布,但较为零散.
此外,当前细枝柃的适生区分布格局与柃木相似,主要集中于华东、华中、华南等地区. 细枝柃的适生区总面积约为1.79×106 km2,其中高适生区占32.3%,中适生区占33.6%,低适生区占34.1%(表 3). 在未来气候情景下,细枝柃的潜在适生区总面积呈增大趋势,分布呈现南北分化,集中于西南、西北和东北地区,主要包括西藏、四川、云南、黑龙江、内蒙古等地. 高适生区在2080-2100年SSP585情景下达到最大,为1.25×106 km2(图 5).
-
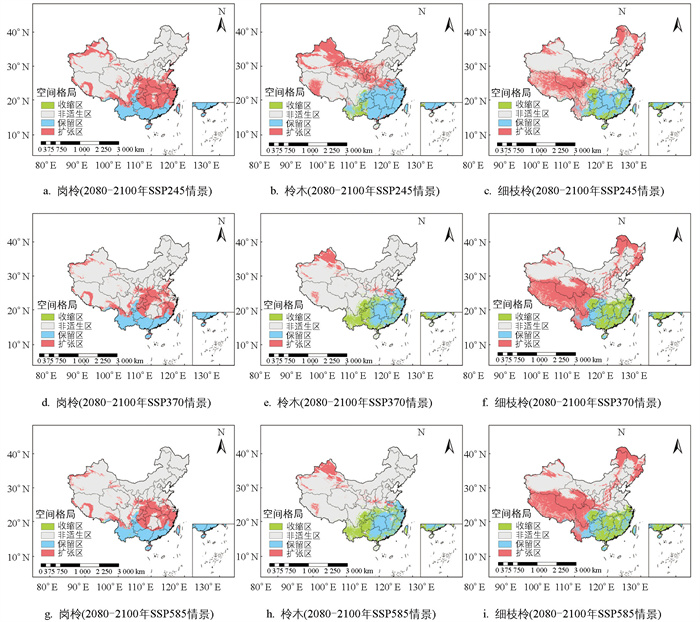
对比分析了2080-2100年SSP245,SSP370,SSP585气候情景下3种柃属植物的潜在适生区空间格局与当前适生区(图 6),结果显示,未来这3种气候情景下,岗柃的潜在适生区总面积大幅扩增,收缩面积极少(表 4). 其中,在2080-2100年SSP245情景下,扩张面积最大,扩张率达72.2%,主要向北转移,但转移距离较短. 当前气候适宜分布区保留率为27.6%,收缩率仅为0.1%(图 6、表 4). 在2080-2100年SSP245情景下,柃木的适生区扩张率达52.4%,主要向西北部地区转移,扩张距离较远(图 6),其适生区保留率为41.1%,高于岗柃. 在2080-2100年的SSP370和SSP585气候情景下,柃木的扩张幅度明显减小,收缩面积增加,尤其是西南地区的分布面积逐渐减少. 气候变化对细枝柃影响显著,其潜在适生区在3种未来气候情景下均明显扩张,主要向西部和东北部转移,其中在2080-2100年SSP585情景下,扩张率达到65.9%. 细枝柃的当前适生区保留率较低,收缩率较高,主要发生在华东和华南地区(图 6、表 4).
2.1. 3种柃属植物的MaxEnt模型精度评估
2.2. 3种柃属植物潜在适生区的重要环境因子贡献率及敏感性分析
2.3. 当前和未来气候情景下潜在适生区预测结果
2.4. 物种的潜在适生区在未来气候情景下的空间格局变化
-
物种的潜在分布区由多种环境因子共同作用,也反映了植物对气候变化的响应[34]. 本研究结果表明,3种柃属植物的主导环境因子是气候变量,其中温度和降水贡献率最高. 具体而言,岗柃和细枝柃的分布格局受最干月降水量的影响,这与前人对濒危植物闽楠分布的研究结果一致[36]. 当最干月降水量在50~150 mm时,岗柃与细枝柃的适生概率较大,且细枝柃的水分需求比岗柃更高,当前气候条件下的栖息地分布图证实了这一结果. 细枝柃主要分布于华中和东南地区,即使在最干月也有充足的降水,而岗柃则主要分布在西南地区,抗旱性较强,可能与东南季风沿海登陆后水汽在向内陆输送过程中逐渐损耗有关[41]. 此外,岗柃和细枝柃为喜光植物,耐阴性较差,常见于日照充足的山坡谷地或林下开阔处[42-43]. 根据响应曲线结果,即使在最冷月,岗柃和细枝柃的适宜生长温度也在25 ℃以上. 因此,温度对柃木分布的影响更大,平均气温日较差越小,柃木的适生概率越高. 树木的径向生长主要发生在夜间,较高的夜间温度使树木保持较高的代谢率,能积累足够的能量和资源来促进树木生长. 较低的昼夜温差还能增加叶片的呼吸作用,提升光合作用速率[44-45]. 此外,柃木的水分需求高于岗柃和细枝柃,因此最湿季降雨量是影响其分布的重要环境因子之一. 高大海等[46]的研究发现,柃木抗旱能力较差,水分充足的地区更适宜其生长.
研究表明,海拔、坡度、坡向等地形因素以及土壤理化性质对植物的生存与分布有显著影响[7, 47-48]. 本研究发现,尽管地形因子和土壤因子未在岗柃和柃木的分布中起主导作用,但土壤pH值对细枝柃的分布有一定调控作用. 具体而言,细枝柃在土壤pH值为5~7时生长良好. 一般在碱性条件下,土壤中的NH4+会转化为NH3并挥发,进而降低有机质和氮的有效性. 因此,常绿和落叶灌木更适宜生长在酸性土壤中[49],而在弱酸性条件下,植物根系对土壤养分的利用率最高[50-52]. 由此可见,土壤和地形环境因子通过直接或间接方式对植物的生长产生重要影响.
-
在2080-2100年不同气候胁迫情景下,3种柃属植物的分布格局发生变化,并呈现向北迁移的趋势. 在2080-2100年SSP245情景下,岗柃与柃木的适生区面积最大,而细枝柃在SSP585情景下适生区面积最大. 陆生植物的分布前沿主要受低温制约[53]. 从2080-2100年SSP245(低胁迫情景,2.97 ℃)到SSP585(高胁迫情景,5.62 ℃),随着未来中国地区温度持续上升[54],温度限制的解除将促进柃属植物向高纬度地区扩展. 其中,岗柃在3种未来气候情景下的保留率和扩张率最高,适生区收缩率不到1%. 此外,高大海等[46]和李世友等[55]的研究发现,岗柃具有较强的抗旱和阻燃能力,抗逆性强,能够有效抵御不利环境变化,可能成为高度胁迫条件下的受益者. 此外,柃木受气候变化影响显著,未来可能向中国西北地区迁移,而现有的西南地区栖息地将大面积收缩. You等[54]在对CMIP6模型年平均温度变化的分析中表明,中国西北部和青藏高原未来温度上升更快,符合柃木生长发育的温度要求. 然而,西北地区属于干旱或半干旱区,2080-2100年在SSP370和SSP585气候情景下,温度大幅上升将加剧环境退化和荒漠化,不利于柃木生存[56]. 此外,我国西南地区多山地高原,在温度升高和雪线上移的影响下[34],低海拔地区常形成干热河谷[57],即高海拔地区气候寒冷,而海拔较低的山谷地区气候干热. 研究表明,柃木的抗旱能力相对较差[46],因此在西南地区生存困难,适生区面积减少. IPCC报告指出,气候变化将加剧水循环,引发更大的降水变化,并导致海平面进一步抬升,我国沿海地区将面临更多的强降雨和洪水[2-3]. 然而,细枝柃的水分需求较低,最干月降水量仅需50 mm便能达到最佳生长状态. 过量水分可能影响植株地下根系的生长,阻碍地下矿质元素的吸收和地上部分的营养供应[58]. 因此,本研究的预测模型显示,细枝柃的潜在适生区在未来将向西部和东北部两极化扩张,而东部沿海地区的栖息地急剧收缩. 尽管在低浓度排放情景下(2080-2100年SSP245气候情景),3种柃属植物可能适应一定范围的温度和降水,其潜在适生区面积也将扩大. 然而,在高浓度排放情景下(2080-2100年SSP585气候情景),持续上升的温度可能超过柃属植物的生态耐受极限[59],对其生存造成不利影响,压缩生存空间.
-
物种分布模型(SDMS)的可靠性取决于以下几个方面[36]. 首先,取决于模型的选择及参数优化. 本研究采用了MaxEnt模型来预测3种柃属植物的潜在适生区. MaxEnt模型源于统计力学,通过找到最大熵的概率分布来估计物种的概率分布. Phillips等[19]认为,MaxEnt能够处理连续变量和分类变量,并考虑不同变量之间的相互作用,从而解释环境变量与物种适宜性之间的关系;此外,MaxEnt模型的概率分布依赖于目标物种的现有分布点,这在很大程度上避免了抽样偏差. 其次,AUC作为模型精度评价指标,其值通过调试模型参数获得,同样影响模型的可靠性. 本研究建立了适用于3种柃属植物的最优预测模型,其AUC值均大于0.9,表明模型预测精度较高. 最后,环境因子的数量、种类及精度也对模型的准确性至关重要. 本研究选取了19个生物气候因子、3个地形因子和6个土壤因子,共计28个环境因子,预测变量范围广泛,分辨率为5×5 km2,精度较高. 因此,本研究对3种柃属植物潜在适生区的预测具有较高的可靠性和可信度. 然而,即使模型已考虑物种的全部生态位需求,预测的潜在适生区通常仍会超过其实际分布[19]. 实际的物种地理分布受到多种因素的制约,如扩散地理限制,生物间相互作用(如捕食、竞争等)和人类活动[60-61]. 其中,人类活动对物种的生存和生长影响日益深远. 例如,曹倩等[62]对青藏高原特有植物祁连獐牙菜进行物种分布建模时,量化了人类活动的影响,包括土地利用、人口密度、建筑设施、道路和夜间灯光等因素. 此外,森林砍伐、放牧和焚烧等人为干扰因素也会导致植物的实际地理分布偏离预测. 因此,未来研究若将更为全面的生物和非生物因素纳入模型中,可能会提高预测的准确性. 尽管如此,本研究准确预测了未来气候变化条件下岗柃、柃木和细枝柃在中国潜在适生区的空间格局变化,为森林资源管理及蜜源植物培育等提供了理论依据.
-
气候是影响3种柃属植物潜在适生区的主要因素. 岗柃和细枝柃的分布主要受最干月降水量驱动,而柃木则主要受平均气温日较差的影响. 在当前的气候条件下,岗柃主要分布在华南地区,尤其是西南部;柃木和细枝柃主要分布在华中和东南沿海地区. 在2080-2100年3种气候情景下,3种柃属植物均呈现向北迁移的趋势,其中,SSP245情景下,岗柃和柃木的潜在适生区扩张最大,而在SSP585情景下,细枝柃的适生区扩张最大. 岗柃在未来长期内具有最高的稳定性,可能成为3种柃属植物中受益最大者.