



 下载:
下载:

-
近百年来,植物物候及其影响因素一直都是生态学家和植物学家关注的焦点[1],其中开花物候是植物物候研究的一个关键,它是植物繁殖对长期自然选择的适应结果,反映了植物周期性生命现象的时序规律对气候等环境因子的响应[2],受到了广泛关注[3].
植物达到一定年龄和体积时就获得了开花能力,进而可通过自主途径或适宜的环境条件诱导而开花.很多研究表明,植物的开花物候由生物因素[4]或/和非生物因素[5]决定.由于乔木与灌木属不同生长型植物,它们在自然界植物群落中处于不同高度,属于群落垂直结构中不同层片,其周边微生境的光照、温度、水分、传粉生物种类和数量等关键生态因子有着显著不同,对生态系统的结构和功能的贡献也有巨大差别.因此,作为自然植物在城市背景下的特殊群体,不同的生长型园林植物开花物候也会存在差异.
事实上,很多研究已经证实在热带和温带地区,不同的开花物候模式与不同的生长型(乔木、灌木、攀援植物)密切相关.例如,一些对热带季节群落的研究发现,大多数木本植物和草本植物分别在不同的季节开花结果[6-7];有的学者观察到,在一些热带森林中不同生长型的植物花期长短也不同[8];也有学者在中国南方研究发现不同的生长型会导致植物的盛花期错位[9].但是这些研究主要集中在热带或温带森林的开花物候及其影响因素等方面[10],对于亚热带地区的园林植物的开花模式的研究还不够充分;同时,关于生长型与花的形态特征的关系的研究还较少,大部分研究都集中在环境因素、遗传因素或授粉者对花的形态特征的影响方面[11-12].
对园林植物开花物候和花形态特征的研究对人类生活和健康有重要意义.很多园林植物的花在发育过程中,具有较高的观赏特性,能为当地带来极大的旅游收益,例如日本的樱花节[13],这是一个具有重要文化意义的节日,每年都有数百万来自海外的游客到日本观赏樱花[14],对园林植物开花物候和花的形态特征的研究可为相应旅游活动的开展提供理论基础[15].除此之外,园林植物开花物候的变化还可以对生态和生产造成强烈的影响,并通过改变暴露于致敏花粉的方式进一步影响人类健康[16].
中国亚热带地区是东亚植物区系多样性的中心,拥有2 600多个属的约14万种种子植物,其中许多种是特有的,具有很大的研究价值.因此,对中国亚热带地区的园林植物的开花物候以及花的形态特征进行研究是相当必要的.本研究选取重庆市主城区常见的木本园林植物作为研究对象,对19个科46种具有不同生长型的木本园林植物进行了为期一年半的观测,记录了每种植物的始花期、盛花期和开花持续时间,并收集了相关物种花朵直径与花朵颜色等数据.目的是研究不同生长型的木本园林植物生长型与其开花物候的生态关系,并探讨其生态关联与花形态特征的可能机制,以及对园林植物配置和管理的实践意义.
全文HTML
-
本研究的研究区域为重庆市主城区(29.35°N,106.33°E),位于中国西南部、长江上游地区,属于青藏高原与长江中下游平原的过渡地带.地势由南北向长江河谷逐级降低,西北部和中部以丘陵、低山为主,北临大巴山、东南部靠武陵山,坡地较多,有“山城”之称.
重庆属亚热带季风性湿润气候,年平均气温为16~18 ℃.年平均降水量较丰富,大部分地区在1 000~1 350 mm,降水多集中在5-9月.年平均相对湿度多在70%~80%之间,在中国属高湿区.年日照时数为1 000~1 400 h,日照百分率仅为25%~35%,为中国年日照很少的地区之一.
-
本研究的观测对象为重庆市主城区常见木本观花园林植物,共计19科、35属、46种(表 1).将所有的观测对象划分为乔木与灌木两种,由于本研究的对象是城市背景下的园林植物而不是自然植物,所以划分乔木与灌木的主要依据是植物的高度,故而将木质藤本植物也记为灌木.此外,对于一些可能具有不同生长型的植物,本研究按照它们的个体实际生长情况进行归类.
观测所选的树种都是发育正常的中龄树,且开花结实3年以上,每种选3~5株作为观测目标.对选定的树木保持其正常生长和发育.所选的观测点,都具有一定的代表性,可以代表那个地方的环境条件(如地形、土壤和植被等),尽量选择平坦、开阔的地方.观测时间为20180101-20190701,涵盖了所有观测树木的完整生命周期.时间记录以“日-旬-季-年”为物候数据采集及数据分析的时间单位体系.时间记录以年日为主,如“20180103”的即记录为“3”.
-
参照“中国物候观测网”介绍的物候观测方法,将植物的花期分3类.始花期:在选定观测的同种树株树上,见到一半以上植株有5%的花瓣完全展开时为始花期.盛花期:在观测树上见有一半以上的花蕾都展开花瓣或一半以上花序松散下垂或散粉时为盛花期.末花期:在观测树上残留约5%的花时为末花期,以花序脱落时为准.春、秋季植物生长发育迅速,至少2 d观测1次;夏季至少3 d观测1次;冬季可7 d观测2次;观测植物物候现象的时间一般在下午,因为一天之内,下午13:00-14:00左右气温最高,而植物的物候现象常在高温之后出现.每次观测时,将观测到的数据记录于制定的“物候观测表”上.
-
为检验不同生长型的木本园林植物开花物候和花形态的关系,首先将同种植物所取的重复样本所记录的观测数据计算出平均值,建立数据库;然后对数据进行Pearson相关性分析和单因素方差分析.所有数据分析利用了SPSS软件(23.0版本).
1.1. 研究区域
1.2. 物候观测对象及时间
1.3. 物候观测记录方法
1.4. 数据分析
-
所有观花园林植物的始花期差异很大,从1月至12月均有植物开花.在乔木中,开花最早的是梅,初花平均年日为全年第4 d,即1月上旬;最晚开花的是木犀与木芙蓉,初花平均年日为全年第244 d,即9月上旬.在灌木中,开花最早的植物是野迎春,初花平均年日为全年第43 d,即2月中旬;开花最晚的是蜡梅,初花平均年日为全年第344 d,即12月中旬.乔木的初花平均年日是全年第97 d,即4月上旬,灌木的初花平均年日为全年第130 d,即5月中旬(图 1).Pearson相关分析发现,生长型与始花期的相关性并不具有统计学意义(p=0.358).也就意味着不同生长型并没有导致始花期的区别.
-
开花持续时间相差很大,介于30 d和306 d之间.在乔木中,开花持续时间最长的是白兰,持续244 d,从4月上旬开至11月下旬;开花持续时间最短的是木犀,仅持续30 d,花期为整个9月.在灌木中,开花持续时间最长的是三角梅,持续306 d,从3月上旬开至12月下旬.开花持续时间最短的有5种,均持续60 d,分别是:紫荆,花期从3月上旬至4月下旬;紫藤,花期从4月上旬至5月下旬;夏鹃,花期从5月上旬至6月下旬;栀子,花期从6月上旬至7月下旬;大花绣球,花期从8月上旬至9月下旬.乔木平均开花持续时间为80 d,灌木平均开花持续时间为131 d(图 2).Pearson相关分析发现,植物的生长型与开花持续时间的相关性具有统计学意义(p=0.010).
-
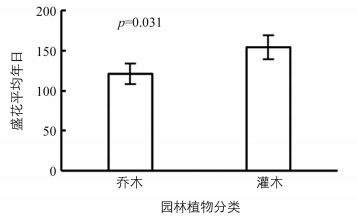
所有观花植物的盛花年日相差很大,从全年第5 d至全年第334 d.盛花年日最早的是蜡梅,为全年第5 d,但是与其他同生长型植物的盛花年日相差太远.本研究认为蜡梅的盛花年日不具有代表性且容易造成统计上的偏差,因此不选择它作为代表.在乔木中,盛花年日最早的是梅,它的盛花年日为第20 d,盛花期为1月中旬至1月下旬,盛花期持续20 d;盛花年日最晚的是木芙蓉,为全年第278 d,盛花期为9月中旬至10月下旬,盛花期持续50 d.在灌木中,盛花年日最早的是紫荆,盛花年日为第79 d,盛花期为3月中旬至3月下旬,盛花期持续20 d;盛花年日最晚的是朱缨花,为全年第334 d,盛花期为11月下旬至12月上旬,盛花期持续20 d.乔木的盛花平均年日为全年第121 d,灌木的盛花平均年日为全年第154 d(图 3).Pearson相关分析发现,生长型与盛花年日之间相关性具有统计学意义(p=0.031).
-
在乔木中,花朵直径最大的是荷花玉兰,直径为17.5cm,花朵直径最小的是木犀,仅有0.45cm;在灌木中,花朵直径最大的是三角梅与木槿,为5.5cm,花朵直径最小的是朱缨花,仅有0.4cm.乔木的花朵直径平均值为5.4cm,灌木的花朵直径平均值为3.5cm.
在所观察到的观花植物中,花色共有6种颜色,分别是白色(6种)、红色(23种)、橙色(1种)、黄色(8种)、蓝色(1种)、紫色(7种).
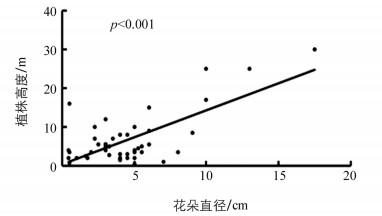
研究发现,生长型与花朵颜色之间相关性不具有统计学意义(p=0.069),而生长型与花朵直径之间的相关性属于边缘具有统计学意义(p=0.059).为进一步探究其相关性,特别对株高与花朵直径之间进行了相关性分析(因为在自然界植物群落中,成年乔木与灌木通常处于不同高度).结果显示株高与花朵直径之间的相关性具有统计学意义(p<0.001),且成正相关关系(图 4).
2.1. 始花期物候特征
2.2. 开花持续时间物候特征
2.3. 盛花期物候特征
2.4. 花形态特征
-
研究发现不同生长型的园林植物与开花持续时间之间的相关性具有统计学意义,并且灌木花期长于乔木花期,这与一些在热带和新热带温带森林中观察到的开花模式相同[8].这可能是因为生长型与植物的生态策略紧密相关,而植物的生态策略可以转化为不同的开花物候行为,所以开花物候会因为生长型的不同而不同[17].而造成这一联系的原因有很多,很多研究表明,开花物候会受到非生物因素(如温度、日照长度和土壤含水量等)或生物因素(如授粉者、生物竞争和系统发育限制等)的影响[18],而不同生长型的植物受到这些非生物因素和生物因素的影响都可能有所不同.
为什么在重庆观察到的观花园林植物中,乔木的花期普遍比灌木的花期短?主要有3个原因.
1) 传粉媒介的差异引发.在园林植物中,乔木的高度普遍高于灌木,在群落中一般处于上层,因此乔木能够更好地利用风作为传粉媒介[19];而灌木一般比乔木矮,且位于群落下层,受风的影响较小,灌木的高度也与大部分传粉昆虫的飞行高度一致,所以灌木更多地利用昆虫作为传粉媒介.一些在热带森林中的研究发现,虫媒物种中观察到的开花期比风媒物种长[8],可能是由于在生物传粉的物种中,这一过程取决于生物的时间和不可预测的行为,所以必须要延长始花期,来弥补这种不确定性,这可以解释为一种对冲策略[20];而风媒传粉物种可以借助风力,能在短时间内同步释放大量花粉,这种授粉方式可能具有优势,因为这增加了受精的可能性.所以风媒物种的花期即使较短,也可以获得较高的传播花粉的机会[21].基于此,有理由相信授粉方式的不同是导致乔木和灌木花期长短不同的主要原因之一.
然而,这样的猜想与植物学的一个普遍认同观点相冲突,即在植物学中普遍认为同一物种的传粉媒介一般相对固定,不会因为生长型的变化而发生变化.本研究提出了一些猜想和假设(如园林植物的特殊性、统计的差异等),但是目前并没有有力的证据来证明这些猜想和假设,所以希望留待后续进一步研究.
2) 应对害虫的策略.乔木由于高度的原因,可能受到害虫伤害的几率比灌木小.有研究证实害虫在花期的长短上也可能有一定的作用,开花物候受到季节变化的生物和非生物因素的选择,而害虫是生物因素中很重要的一员[22].某些植物为了补偿特定的害虫对其繁殖的伤害,可能选择延长自己的花期来弥补这种伤害.植物不能同时既避免授粉不足、传播不足、发生病害又能够传播大量的花粉,在这些风险之中需要进行权衡,而灌木延长开花期可能是一种更好的对冲策略,可以在足够的花粉传播和被害虫伤害之间进行权衡[23].
3) 日照长度效应.在重庆主城区的园林植物景观中,乔木、灌木和草本植物一般都是配合使用,而由于灌木的高度普遍低于乔木,导致灌木一般处于群落的下层,受到的日照时间更短,所以灌木的花期更长.其中可能的机制是,除生物因素外,非生物因素也对不同生长型的植物的花期长短有影响.例如,一些研究显示日照长度和植物的花期长短成反比[8];也有研究发现相对于冠层植物,林下物种的花期更长[24].但是关于为什么日照长短与开花物候成反比,是否有什么内在的生理生化因素,还有待于进一步研究.
-
据观察,园林植物的生长型与盛花期之间相关性具有统计学意义,乔木盛花平均年日为第121 d(5月上旬),灌木盛花平均年日为第154 d(6月上旬),灌木盛花平均年日比乔木盛花平均年日晚约30 d.
在一些研究中也显示了相似的结果[9],但是并没有明确的解释.有的学者认为这是由系统发育的限制造成的[25],也有学者认为温度的季节性变化是对植物开花模式影响最大的环境因素[26].然而,在亚热带地区,温度的季节性变化并没有温带地区那么强烈,特别是在重庆,冬季最冷的时候温度也很少低于摄氏零度,所以显然温度不是造成这一现象的主要原因.
我们认为,降水量和授粉者可能才是导致这一结果的主要因素.首先,重庆地区降水量的特点是在雨季中6月降水量普遍到达最高峰,接近500 mL,而5月上旬刚进入雨季,降水量并不大,只有200 mL左右.另外,不同生长形式的植物开花不仅与降雨量密切相关,并且在可能的授粉载体方面也有差异.对授粉者的研究显示,授粉者通常只有在一定的个体或花朵临界密度开花后才会被特定的植物物种所吸引[27],而开花的时间分布与花授粉的合适的生物和非生物条件密切相关,这影响了物种的成功繁殖[20].所以,由于降雨量对花粉传播的影响,导致不同生长型的植物在雨季进化出了不同的盛花期.因为乔木普遍位于冠层,而且授粉方式以风媒为主,受降雨影响较大,所以在雨量最大的前一个月开花达到高峰期,以避免6月高强度降雨对花粉传播的影响.而灌木位于林下,由于乔木的遮蔽,其受降雨的影响较小,而且为了适应授粉昆虫季节性的活动,故选择在6月中旬达到盛花期.
-
生长型与花朵直径之间的相关性虽然属于边缘具有统计学意义,但是植株高度与花朵直径之间相关性极具有统计学意义且为正相关关系.因为在自然界中,成熟的乔木普遍高于灌木,所以这一结果可以表明乔木的花朵直径一般大于灌木的花朵直径,即不同的生长型导致了不同大小的花朵直径.
关于花朵直径的影响因素,很多研究者认为,植物花朵的最终大小是由生物因素与非生物因素共同作用的结果[28].例如一项在中国西部高海拔地区的研究显示,随着海拔的升高和温度的降低,狼毒花的花朵直径逐渐增大[29];而另一项关于菊花的研究表明,在其他条件不变的情况下,适当增大光照强度或降低温度都能使菊花长出更大的花朵[30];同时,在很多研究中都观察到对传粉者的吸引力的大小在虫媒植物的繁殖中起着决定性的作用[31],更大的花能吸引更多的传粉者[32].
由于乔木高于灌木位于群落的较高层片,所以乔木所处的层片的温度普遍比灌木所处的层片的温度低并且受到更多的太阳辐射,而与此同时,由于温度降低和辐射增强导致授粉者减少,所以异花传粉植物必须要适当增大花朵来吸引传粉者.所以温度、光照强度以及对授粉者的吸引可能是导致这一差异的主要原因.
在研究中还发现,花色与生长型的相关性并不明显,植株高度与生长型之间也没有具有统计学意义的相关性,这是一个奇怪的现象,因为在很多研究中都表明花色与植株的高度和所处的环境有着密切的关系[33],但是本研究并未观察到这一点.考虑到本研究仅仅研究了重庆市主城区常见的木本观花园林植物,只占重庆市所有木本植物的极小部分,我们认为可能是样本选择的偶然性导致了这一结果.
-
本研究发现始花期与生长型的相关性并不具有统计学意义.从表面上看,灌木的平均始花期比乔木晚大约1个月,貌似因为不同的生长型导致了不同的始花期,但是数据分析发现,生长型与始花期之间的相关性并不具有统计学意义,说明它们之间并没有明显的联系,统计数据的差异很可能是记录方式导致的结果.
-
为了探讨植物原产地对研究内容所带来的影响,分析了植物原产地的气候条件与开花物候之间的相关性.研究发现,始花期、盛花期与植物原产地的气候条件均不具有统计学意义相关性,而开花持续时间与原产地的气候条件之间相关性具有统计学意义(p=0.010),具体表现为随着植物原产地气温的升高,开花持续时间相应增加.这表明,除生长型外,植物原产地的气候条件也可能是影响开花持续时间的一个重要因素.
3.1. 开花持续时间与生长型
3.2. 盛花期与生长型
3.3. 花形态特征与生长型
3.4. 始花期与生长型
3.5. 原产地与开花物候
-
本研究证实在位于亚热带季风气候的重庆地区,园林植物的盛花期、开花持续时间与植物的生长型密切相关,但是始花期并未受到不同生长型的影响.具体而言,是生物因素与非生物因素共同导致了这一结果,其形成可能机制与花的大小有关,也可能由于传粉媒介、应对害虫的策略、对日照长度的反应等方面的不同所导致,但与花朵颜色无关.对亚热带地区观花园林植物的开花物候与花朵的形态特征的研究,在理解该类型地区影响植物功能性状的因素方面有所启迪,相信本研究的结论和产生的信息能够作为未来进一步研究的基础或是景观设计与旅游规划的重要参考.
此外,本研究也有一些不足之处.如本研究的植物仅为46种木本观花园林植物,这只占重庆市的所有木本植物的一小部分;本研究在尺度上仅限于内陆的亚热带季风气候区域,涉及研究范围较小.这两点可能限制了研究结果的普适性.对于一些猜想与假设,如不同生长型木本园林植物的传粉媒介具有差异这一点,本研究并不能提出有力的证据,希望在后续的进一步研究中能有所进展.