


 下载:
下载:
-
开放科学(资源服务)标志码(OSID):

-
土壤碳(C)、氮(N)、磷(P)是植物生长所必需的营养元素,在生态系统的演替过程中起重要作用[1]. 土地利用是影响土壤C,N,P循环最主要的因素之一[2]. 不同土地利用方式下植被类型、养分输入、管理措施、侵蚀强度及理化性质等不同,显著改变了土壤环境,进而导致土壤C,N,P质量分数及其生态化学计量比发生改变[3-6]. 土壤C,N,P生态化学计量影响着植物初级生产力、微生物活动、土壤质量等,是全球生物化学平衡与循环的关键因素[7-9]. 因此,研究不同土地利用下土壤C,N,P的生态计量特征对揭示土壤C,N,P有效性及其平衡与循环机制具有重大意义[10].
紫色土是四川盆地一种典型的土壤类型,其矿质养分丰富,肥力较高,农业利用价值很高. 然而,由于降雨丰富,农业生产活动频繁,垦殖率高,紫色土区的水土流失较为严重[11]. 近年来,为取得较好的水土保持效益,紫色土部分区域实施了耕地转林地、园地等水土流失治理措施[12-13]. 这些措施改变了土地利用方式,导致土壤C,N,P质量分数及生态化学计量比发生变化,但其变化程度和量级尚未量化[14-15]. 鉴于此,本研究通过对比分析同一紫色土区坡面3种不同土地利用方式(耕地、园地、林地)下土壤C,N,P质量分数及其生态化学计量比差异,揭示不同土地利用方式对土壤生态化学计量变化的影响特征,为区域土地的合理利用及生态恢复重建提供科学指导.
全文HTML
-
研究地点位于重庆市忠县石宝寨石盘丘小流域(108°10′25″E,30°24′53″N),其地势南低北高,海拔119~780 m. 该区域气候为典型湿润季风气候,年均降雨量1 100 mm,年均气温为18.5 ℃. 该小流域总面积约0.44 km2,土壤以中性紫色土和水稻土为主,主要土地利用方式为耕地(35.46%)、林地(28.17%)、园地(13.78%). 耕地主要为玉米、油菜轮作,每年施尿素250~350 kg/hm2和标准复混肥300~600 kg/hm2;园地树种为柑橘,每年施标准复混肥500~900 kg/hm2;林地树种主要以马尾松为主,夹杂柏木、栎属和杉木等,无施肥.
-
2019年5月分别在3种土地利用方式的(耕地、园地、林地)坡顶、坡中、坡脚各选择3个代表性样地,每个样地面积为10 m×10 m. 按“S”型布设7个采样点,各点均采集0~5,5~10,10~15,15~20 cm土层样品,并将相同土层7个采样点的土壤均匀混合后,用四分法收集1 kg混合土样,装入塑料袋编号后带回实验室备用. 同时,各土层7个采样点均用铝盒和环刀采样,密封后带回实验室测定土壤含水率(SMC)和容质量(BD).
实验室内,将混合土样自然风干后去除砾石、植物等杂物,研磨过筛(1.0 mm),分成2份. 一份用于测定土壤pH值、电导率(EC)和阳离子交换量(CEC),另一份进一步筛分(0.25 mm)后用于测定土壤有机碳(C)、全氮(N)和全磷(P)质量分数. 各指标的具体测定方法为:C采用重铬酸钾外加热法,N采用半微量开氏蒸馏法,P采用钼蓝比色法. 土壤含水率(SMC)采用烘干法,土壤容质量(BD)采用环刀法,pH值采用电位法,土壤EC,CEC分别采用电导法、醋酸铵法[16].
-
采用SPSS 18.0软件进行数据处理和分析. 采用单因素方差(One-way ANOVA)和Turkey法比较不同土地利用方式间及相同土地利用方式各土层间土壤C,N,P质量分数及化学计量比的差异性. 采用Pearson系数表征C,N,P质量分数及生态化学计量比与其他土壤性质(SWC,BD,pH值,EC,CEC)之间的相关性. 显著性水平为:p<0.05为差异有统计学意义,p<0.01为差异极有统计学意义. 采用Originlab 2018绘图. 本研究中土壤C,N,P的生态化学计量比均为摩尔比.
1.1. 试验区概况
1.2. 样品采集与分析
1.3. 数据处理
-
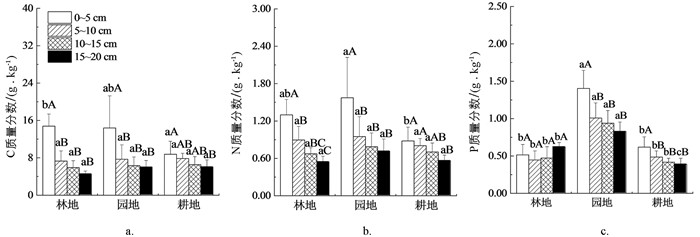
如图 1所示,0~5 cm土层中,园地C质量分数与林地、耕地差异无统计学意义(p>0.05),而林地C质量分数显著高于耕地(p<0.05);5~10,10~15,15~20 cm土层中,不同土地利用之间C质量分数差异均无统计学意义(p>0.05). 0~5 cm土层中,林地N质量分数与园地、耕地差异无统计学意义(p>0.05),但园地N质量分数显著高于耕地(p<0.05);5~10,10~15,15~20 cm土层中,各土地利用之间N质量分数差异均无统计学意义(p>0.05). 0~5,5~10,10~15土层中,园地P质量分数显著高于林地、耕地(p<0.05),而林地与耕地差异无统计学意义(p>0.05);15~20 cm土层中,各土地利用之间P质量分数差异均有统计学意义(p<0.05),从大到小依次为:园地(0.83 g/kg)、林地(0.62 g/kg)、耕地(0.39 g/kg).
林地中,0~5 cm土层C质量分数显著高于其他土层(p<0.05),其他土层间C质量分数差异无统计学意义(p>0.05);0~5 cm土层N质量分数显著高于其他土层,且5~10 cm土层N质量分数也显著高于15~20 cm土层(p<0.05);各土层间P质量分数差异无统计学意义(p>0.05). 在园地、耕地中,0~5 cm土层的C,N,P质量分数均显著高于其他土层(p<0.05),而5~10,10~15,15~20 cm土层之间差异均无统计学意义(p>0.05).
-
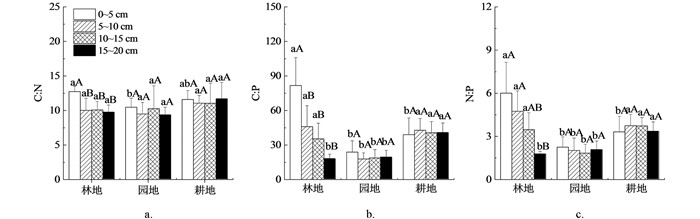
如图 2所示,0~5 cm土层中,耕地C∶N与林地、园地差异无统计学意义(p>0.05),而林地C∶N显著高于园地(p<0.05);5~10,10~15,15~20 cm土层中,不同土地利用方式之间C∶N差异无统计学意义(p>0.05). 0~5 cm土层中,林地C∶P显著高于园地、耕地(p>0.05),而园地与耕地差异无统计学意义(p>0.05);5~10,10~15 cm土层中,园地C∶P显著低于林地、耕地(p>0.05),而林地与耕地差异无统计学意义(p>0.05);15~20 cm土层中,耕地C∶P显著高于林地、园地(p>0.05),而林地与园地差异无统计学意义(p>0.05). 0~5 cm土层中,林地N∶P显著高于园地、耕地(p>0.05),而园地与耕地差异无统计学意义(p>0.05);5~10,10~15 cm土层中,园地N∶P显著低于林地、耕地(p>0.05),而林地和耕地差异无统计学意义(p>0.05);15~20 cm土层中,耕地N∶P显著高于林地、园地(p>0.05),而林地与园地差异无统计学意义(p>0.05).
林地中,0~5 cm土层C∶N,C∶P,N∶P显著高于其他土层(p>0.05),其他土层间C∶N,C∶P,N∶P差异无统计学意义(p>0.05). 在园地、耕地中,各土层之间C∶N,C∶P,N∶P差异均无统计学意义(p>0.05).
-
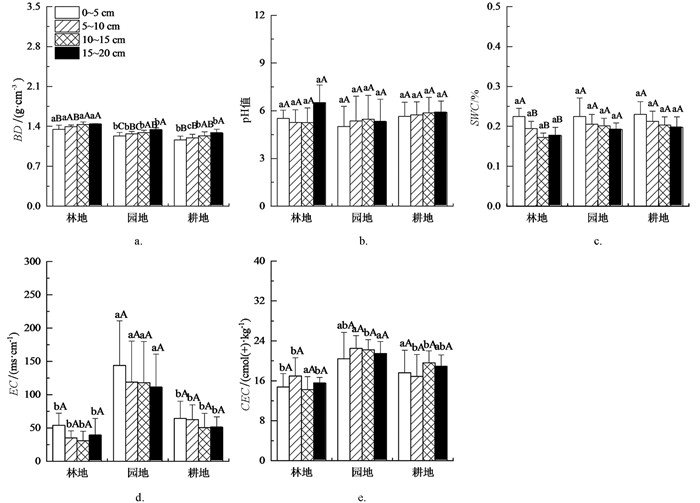
如图 3所示,在所有土层中,林地BD均显著大于园地、耕地(p<0.05),园地与耕地差异无统计学意义(p>0.05);不同土地利用方式间pH值、SMC差异均无统计学意义(p>0.05);园地EC显著高于林地、耕地(p<0.05),林地与耕地差异无统计学意义(p>0.05);不同土地利用方式间CEC差异有统计学意义(p<0.05).
同一土地利用方式中,BD,pH值随土层递增,SMC,EC随土层加深递减. 在林地、园地、耕地中,15~20 cm土层BD显著大于0~5 cm土层(p<0.05);各土层之间pH值,SMC,EC,CEC差异均无统计学意义(p>0.05).
-
由表 1可看出,土壤C,N质量分数与BD,pH值呈极显著负相关(p<0.01),与SMC呈极显著正相关(p<0.01),N质量分数与CEC呈显著正相关(p<0.05);土壤P质量分数与EC,CEC呈极显著正相关(p<0.01),C∶N与CEC呈显著负相关(p<0.05);C∶P,N∶P与EC,CEC均呈极显著负相关(p<0.01).
2.1. 土壤C,N,P质量分数
2.2. 土壤C,N,P生态化学计量比
2.3. 其他土壤性质
2.4. 土壤C,N,P生态化学计量比与土壤各因子的相关性
-
本研究结果表明,林地与园地C,N质量分数总体上高于耕地,这与罗由林等[17]研究结果相似. 施肥增加了耕地和园地中土壤C,N输入量,但耕地植被覆盖远低于园地,农业翻耕等活动频繁,破坏了土壤团粒体结构,促进土壤有机质矿化和土壤侵蚀,导致了C,N元素大量流失[4, 15, 18]. 此外,与耕地相比,林地、园地内的植被枯枝落叶能有效增加表层土壤C,N质量分数,加之浅层根系发达,土壤结构良好,有利于土壤C,N固存[19]. 在本研究中,园地土壤P质量分数显著高于林地、耕地. 这可能是因为一方面园地施肥增加了外源P肥输入,另一方面,相比耕地,园地水土流失程度较轻,减少了土壤P流失.
各土地利用方式下,土壤C,N质量分数随土层深度增加而递减,表层土壤C,N质量分数(0~5 cm)显著高于其他土层,这与杜映妮等[20]的研究结果相似. 这可能是因为表层土壤易受外界环境因素的影响,植物的枯枝落叶及施肥均为表层土壤提供了充足的C,N源,呈现出表聚性[21]. 水分垂直运动可使C,N向下层迁移,但在迁移过程中,其扩散能力随土层深度增加而递减,使得下层土壤C,N质量分数增加不明显[22]. 土壤P在耕地和园地中同土壤C,N一样呈表聚性,而在林地15~20 cm土层中最高. 这可能是因为不同土地利用方式下土壤P的主要来源不同. 在耕地、果园等土壤中,因长期施肥导致了土壤P呈现表聚性,而林地土壤P主要来源于土壤母质风化产物,因此林地15~20 cm土层P质量分数最高[23-24].
-
本研究中,土壤C∶N值从大到小依次为:耕地(11.33)、林地(10.72)、园地(9.90),土壤C∶P值从大到小依次为:林地(49.94)、耕地(40.82)、园地(20.02),土壤N∶P值从大到小依次为:林地(4.48)、耕地(3.54)、园地(2.05),表明土地利用方式可导致C,N,P生态化学计量变化,这与王维奇等[25]的研究结果相似. 这可能是因为各土地利用方式下土壤C,N,P的积累与流失机制不同. 一方面,与施肥增加耕地C,N,P输入量相比,林地通过植被反馈积累C,N,P元素,而园地则通过施肥及植被反馈共同积累,不同的输入方式可使土壤C,N,P质量分数呈不同比例增加,而改变它们的生态化学计量比. 另一方面,耕地与林地、园地相比,水土流失更严重[15];此外,土壤C,P主要以颗粒态流失,N主要以溶解态流失[26],土壤C,N,P不同的流失形态使得土壤C,N,P在不同侵蚀强度下流失比率会不一样,这也使得3种土地利用方式下土壤生态化学计量比产生差异. 因此,导致在3种土地利用方式下耕地土壤C∶N高于林地、园地,林地土壤C∶P,N∶P高于耕地. 本研究中,耕地C∶P,N∶P均大于园地,这可能是因为园地土壤P质量分数较高,且能够更好地固存外源输入性P肥,使得园地C∶P,N∶P较低. 不同土地利用方式下,各土层之间C∶N,C∶P,N∶P差异均无统计学意义,表明各土层间C,N,P的比例变化不大,这与Li等[27]的研究结果相似.
土壤C,N,P生态化学计量比是衡量C,N,P元素平衡与循环,判断土壤有机物组成及矿化、累积程度的重要指标[1]. 研究区各土地利用方式土壤平均的C∶N(10.63),C∶P(36.08)及N∶P(3.32)均低于中国土壤平均值(分别为13.9,154.9,11.3)[28]及全球土壤平均值(分别为14.3,186.0,13.1)[29],表明N为该区域土壤限制元素,这与Li等[27]结果相似. 这可能是因为相比于C和P,N更易随径流发生淋溶和横向迁移流失. 降雨径流通常优先搬运土壤中N、极少的可溶性C和P、较轻的植物残体和凋落物,而泥沙开始迁移时,土壤C,P才随泥沙开始大量流失[15, 30].
-
土壤BD,pH值与土壤C,N质量分数皆呈极显著负相关,这与张晗等[31]研究结果一致. 土壤BD反映土壤疏松程度,BD小,土壤下渗及蓄水能力强,有利于植物生长和土壤养分积累[32]. pH值显著影响土壤微生物及酶的活性进而影响土壤C,N固定和积累能力;pH值减小时,微生物及酶分解活动减弱,有利于土壤C,N贮存[33]. 土壤SMC与土壤C,N质量分数呈极显著正相关,这与丁小慧等[34]的研究结果一致. 土壤SMC与土壤养分循环、微生物活动、植物光合生理过程密切相关[35-36];较高的土壤SMC厌氧细菌反硝化作用增强,矿化速率降低,固氮能力增强,有利于C,N矿化积累[37]. 土壤EC,CEC与土壤P质量分数呈极显著正相关,与土壤C∶P,N∶P呈极显著负相关,这与宋佳龄等[38]研究结果相似. 土壤EC与含盐量呈显著相关关系,常用来表示含盐量的高低[39]. 本研究中EC对土壤P的固定有促进作用. 酸性紫色土土壤P固定主要通过铝、铁离子[40],CEC一定程度上可以代表土壤的固P能力. 当CEC增大时,紫色土中铝、铁离子增多,对磷酸根离子的吸附增强,土壤固P能力变强[38, 41].
3.1. 不同土地利用方式下土壤C,N,P质量分数特征
3.2. 不同土地利用方式下土壤C,N,P生态化学计量比特征
3.3. 不同土地利用方式下土壤C,N,P质量分数及生态化学计量比的影响因素
-
土地利用方式显著影响土壤C,N,P质量分数及其生态计量比. 由于林地、园地和耕地3种土地利用方式C,N,P的积累与流失机制差异,耕地土壤C,N,P质量分数最低,但其C∶N相对最大,而C∶P,N∶P低于林地,高于园地. 各土地利用方式下C,N质量分数和耕地、园地P质量分数均为0~5 cm土层最高,而林地P质量分数15~20 cm土层最高(p<0.05);各土层间C∶N,C∶P,N∶P差异也无统计学意义(p>0.05). 土壤C,N质量分数与土壤BD,pH值及SMC相关性有统计学意义(p<0.05),土壤P质量分数,C∶P,N∶P则与EC,CEC相关性有统计学意义(p<0.05). 研究区各土地利用方式下土壤的N∶P均值小于中国及世界土壤的平均值,表明N为该区土壤限制性营养元素.