





-
开放科学(资源服务)标识码(OSID):

-
叶片是植物的重要器官,可通过光合作用为植株提供大量的能量和有机物,以保证其正常生长发育. 叶片衰老作为叶片自然发育的必经阶段,由外部信号和内部信号引起,是一个高度程序化的过程[1]. 该过程中细胞结构、基因表达和细胞代谢有序变化,最初为叶绿体的破裂,接着叶绿素、蛋白质、膜脂、RNA等有机大分子开始分解并向种子或者生长点转移. 该过程还受植物激素、一些代谢物的内源性因素以及光合作用状态等调控[2-4]. 叶片衰老作为植物发育过程中的重要环节,研究叶片衰老在发育生物学中具有积极的意义[5].
水稻是单子叶重要的模式植物,更是人类最重要的粮食作物之一,其增产对世界的粮食安全具有重要意义[6-7]. 籼型品种在中后期叶片易早衰,导致光合能力降低,限制了更多有机物的形成和积累,不利于结实率和千粒质量等产量相关性状的提高[8]. 叶片早衰是指水稻抽穗期至成熟期,叶片内部生理功能失调,生理活动受到限制,导致“未老先衰”的表型,出现茎叶枯萎、失绿及根系活性减弱等症状,致使源器官有机物合成不充足,籽粒不充实,空秕率迅速上升,千粒质量下降,对水稻产量及米质造成极大的损失和影响[9]. 在粮食生产中,水稻叶片早衰现象广泛存在. 有些水稻品种由于叶片和根系早衰而造成结实率偏低、空秕率较高,影响其产量潜力的发挥. 理论上推算,水稻叶片如果推迟1d衰老,可使水稻增产2%左右,而实验结果表明可增产约1%[10]. 有目的地对叶片早衰进行调控,可以提高农作物的产量或延长农产品的贮藏期,因此,开展水稻叶片早衰相关突变体的表型鉴定,以及叶片衰老的生理生化指标检测和基因克隆研究对其在水稻的遗传改良和衰老机制解析等方面具有重要意义.
叶片衰老过程中表达水平发生变化的基因称为叶片衰老相关基因(senescence-associated genes,SAGs). 目前,关于叶片早衰的机理研究较为深入,大多集中在分子和遗传水平上,已有ESL9,OsDOS,OsPLS1,SPL28等20多个调控水稻SAGs完成了基因定位和部分功能解析[11]. 根据早衰突变体表型可分为两种类型:第一个类型是叶片黄化,早衰突变体esl9分蘖期叶尖开始黄化,逐渐延伸至叶片中段,一直持续到成熟期,其调控基因位于第11染色体,引起叶片淀粉积累,使光合系统受阻[12]. OsDOS RNAi植株在孕穗期时叶片开始发黄,灌浆期时叶片明显发黄,OsDOS可能通过或部分通过将发育信号与茉莉酸甲酯信号传导通路进行整合来延缓叶片的衰老[13]. OsAkaGal的过表达植株表现为生长发育延迟且叶片变得灰绿,OsAkaGal是一个类囊体膜降解酶,在水稻叶衰老过程中通过参与双半乳糖甘油二脂的降解从而调控水稻叶片衰老[14-15]. 第二个类型是叶片斑点,早衰突变体ospls1分蘖期下部叶片出现类病斑症状,由叶尖和叶缘扩展至叶片内侧,其调控基因OsPLS1通过参与SA代谢途径引起叶片衰老[9]. SPL28编码1个网格相关受体蛋白复合体亚基(AP1M1),参与调控高尔基体的物质运输途径和囊泡运输,功能缺陷造成水稻过敏性反应,叶片表现出类病斑,导致植株早衰[16]. 突变体rls1早衰的叶片在整个表面形成一些散布的、类似感病的黄棕色小斑点,RLS1编码1个具有NB-ARM结构域的蛋白,参与叶绿体的降解,引发叶片快速衰老[17]. 叶片衰老是一个复杂的过程,尽管部分水稻叶片衰老基因被定位和克隆,但在蛋白合成降解、细胞程序性死亡、激素信号转导等阶段还缺乏系统的认知,需挖掘更多叶片早衰的调控基因,不断丰富叶片早衰的调控网络,为水稻抗早衰品种选育提供坚实的理论基础.
本研究从涪恢9802和LR杂交后代中鉴定了早衰突变体psl4(t),并对该叶片早衰基因进行表型分析和基因定位. 该突变体在6叶期前叶片呈正常绿色,从7叶至剑叶每张叶片从叶尖至叶中部逐渐衰老,其早衰调控候选基因定位于第4染色体上,与已报道的早衰基因位点均为非等位,是一个新的早衰基因. 本研究对该水稻早衰基因的克隆、功能分析及分子调控机理的研究奠定了基础.
HTML
-
水稻早衰突变体psl4(t)是从重庆市渝东南农业科学院以涪恢9802和LR杂交后代中获得的,经连续多代自交选择,早衰特性能够稳定遗传. 之后配制psl4(t)/0739NB,psl4(t)/431B和psl4(t)/中九B等3个杂交组合,利用F1及其自交F2群体进行遗传分析和基因定位. 突变体psl4(t)和野生型(WT)种植于重庆市渝东南农业科学院(重庆涪陵)试验基地,常规肥水管理.
-
水稻成熟后,分别取psl4(t)及WT各20株,对其株高、穗长、有效穗数、每穗粒数、结实率和千粒质量等主要农艺性状进行考种分析. 每株随机抽取20籽粒,利用游标卡尺测定籽粒的长、宽、厚及长宽比.
参考Wellburn[18]的方法,抽穗期测定WT和psl4(t)倒1叶至倒3叶的光合色素质量分数. 准确称样品叶片各0.05 g,并剪成均匀块状,将其放入装满95%乙醇溶液的4 mL离心管中,在4℃条件下避光处理24 h,轻轻摇晃多次,使色素完全溶解. 吸取200 μL上清液,采用酶标仪分别测量665,649和470 nm波长处的吸光值. 光合色素质量分数计算公式参照文献[19].
-
参照He[20]的方法用电子显微镜观察psl4(t)抽穗期剑叶叶尖黄色部分和WT叶尖部的细胞结构. 戊二醛双重固定后,利用不同梯度的乙醇逐级脱水,然后进行置换和包埋,超薄切片后,以醋酸双氧铀和柠檬酸铅液双重染色,荷兰飞利浦TECNAI10电子显微镜观察并照相.
-
大田栽培条件下,调查psl4(t)/0739NB,psl4(t)/431B和psl4(t)/中九B的F1杂种叶色,确定控制突变体叶尖早衰症状的显隐性;根据其F2群体中具有正常叶色的水稻单株数与具有突变体性状的单株数的比例,确定控制叶尖早衰性状的基因数量;随后分单株选取亲本材料psl4(t),中九B,同时在psl4(t)/中九B的F2群体中选取15株具有突变体性状的单株以及15株具有野生型表型的植株叶片,采用CTAB法提取亲本和基因池DNA,碱煮法提取F2代单株总DNA;构建正常基因池和突变基因池,进行连锁标记筛选,引物与PCR反应体系参考王晓雯等[21]的方法. SSR(
http://www.gramene.org/ )分子标记引物由上海生工生物公司合成(表 1),利用BSA法进行基因连锁分析[22]及基因定位,用Mapmaker 3.0软件分析数据并作图,根据Gramene网站(http://www.gramene.org/ )的水稻基因组信息构建物理图谱.
1.1. 试验材料
1.2. 农艺性状及光合色素质量分数的测定
1.3. 透射电子显微镜观察
1.4. 遗传分析及基因定位
-
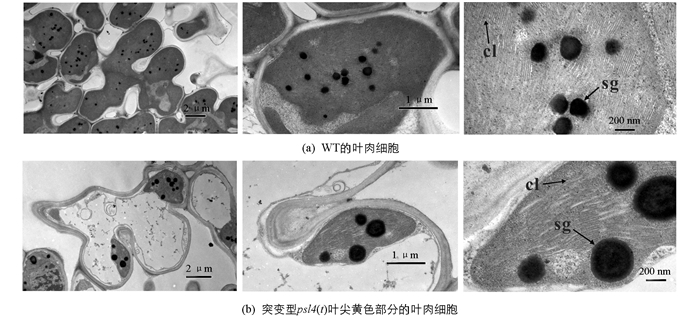
突变体psl4(t) 1~6叶期的叶片呈正常绿色,从7叶至剑叶每张叶片从叶尖至叶中部逐渐衰老. 叶片叶绿体发育受阻其体积变小、光合色素质量分数减少,叶片提前衰老. 抽穗期时,突变体psl4(t)的抽穗期极显著长于野生型(图 1a),突变体psl4(t)的倒4叶出现叶尖早衰黄化(图 1b);成熟期时,突变体psl4(t)主茎长度显著短于野生型(图 1c),所有叶片全部表现严重黄化衰老症状(图 1d). 透射电镜分析表明,分蘖后期至抽穗期突变体psl4(t)剑叶部分叶绿体中的类囊体已开始降解,而且其叶绿体中的淀粉颗粒数量明显少于对照,但淀粉颗粒大小明显增大(图 2). 此外,我们对野生型和突变体psl4(t)的主要农艺性状进行了统计,结果显示突变体psl4(t)的株高、每穗粒数和结实率极显著低于野生型,分别下降8.49%,12.91%和7.73%(表 2),表明突变体psl4(t)叶绿体结构的破坏可导致叶片黄化早衰,影响植株的株高及产量.

-
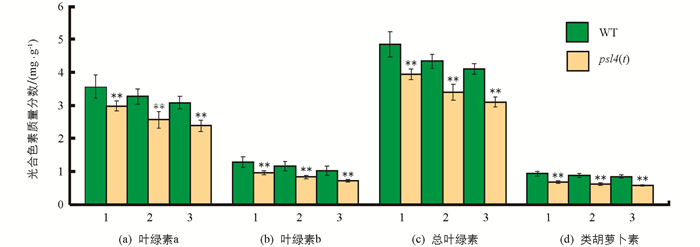
对抽穗期野生型和突变体psl4(t)倒1叶至倒3叶的光合色素质量分数进行测定,结果表明突变体psl4(t)倒1叶(剑叶)、倒2叶和倒3叶的叶绿素a、叶绿素b、总叶绿素及类胡萝卜素的质量分数(图 3)均极显著低于野生型(WT). 与野生型相比,突变体psl4(t)的倒1叶、倒2叶和倒3叶的总叶绿素质量分数分别下降15.37%,14.80%和4.49%. 因此,突变体psl4(t)叶片黄化早衰很可能是光合色素质量分数显著降低所致.
-
突变体psl4(t) 1~6叶期的叶片呈正常绿色,从7叶期至剑叶期每张叶片从叶尖至叶中部逐渐衰老. F1代表现为正常的叶色和寿命,说明该突变性状表现为隐性遗传. psl4(t)/0739NB,psl4(t)/431B和psl4(t)/中九B的3个F2代群体出现正常叶和衰老叶两种情况,正常叶与早衰叶的单株数都符合经典遗传学分离比例3∶1(表 3),表明突变体psl4(t)是由单隐性核基因控制.
-
基因定位中连锁标记的筛选采用近等基因池分析法. 利用psl4(t)/0739NB的F2群体中具有突变性状的102个单株作为基因初步定位群体,选择该群体中15株野生型叶片等量混合构建野生型基因池、15株早衰型叶片等量混合构建早衰型基因池,用均匀分布于12条染色体的500对SSR标记逐条分析其在psl4(t)和0739NB间的多态性,其中106对标记在亲本间表现多态性. 用在亲本间表现多态性的标记对野生型基因池和早衰型基因池进行多态性分析. 结果位于第4染色体长臂的SSR标记RM1155,RM142和RM3524在突变体基因池和正常基因池之间存在明显偏分离,说明psl4(t)位于第4染色体. 利用102个早衰型单株验证以上标记,并根据重组单株数和重组单株分布情况,初步把psl4(t)定位在RM142和RM3524标记之间,遗传距离分别为8.0 cM和10.3 cM(图 4a).
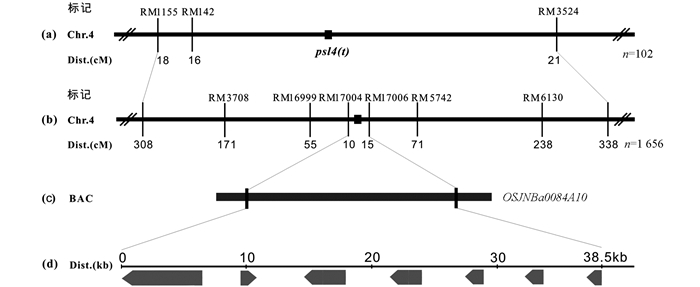
为了将psl4(t)限定在更小的范围内,我们利用RM1155和RM3524标记之间的16对标记,对亲本及基因池进行PCR扩增,找到其中的差异引物RM3708,RM16999,RM17004,RM17006,RM5742和RM6130(表 1). 利用分离群体的1 656个隐性单株将psl4(t)精细定位在BAC克隆OSJNBa0084A10的标记RM17004和RM17006之间38.5 kb的范围内(图 4b和4c). 根据日本晴基因组序列预测在这38.5 kb范围内有7个候选基因(图 4d和表 4),分别是LOC_Os04g35260,LOC_Os04g35270,LOC_Os04g35280,LOC_Os04g35290,LOC_Os04g35300,LOC_Os04g35305,LOC_Os04g35310.
2.1. 突变体psl4(t)的表型及主要农艺性状
2.2. 光合色素质量分数测定
2.3. 突变体psl4(t)的遗传分析
2.4. 突变体psl4(t)的基因定位
-
水稻叶片是光合作用、呼吸作用、蒸腾作用等重要的场所,同时也是影响水稻产量的源器官,因此克隆相关水稻叶片早衰基因对于解析叶片早衰分子机制具有重要意义. 本研究中的突变体psl4(t)属于叶片黄化类型突变体,6叶期前叶片呈正常绿色,从7叶期起至剑叶,叶尖开始黄化沿叶中部逐渐延伸;进一步通过农艺性状统计发现,其穗长、有效穗和籽粒宽变化无统计学意义,而株高、每穗粒数、结实率及千粒质量则显著降低. 早衰突变体psl1自苗期的心叶抽出开始,前1叶就开始变黄衰老[23];psl3则是从第6叶开始表现衰老,叶片从上部到基部衰老程度逐渐减轻,叶下部和叶鞘基本保持绿色[24];pse(t)叶片从抽穗期开始表现衰老,先在老叶上出现褐色斑点,接着整张叶片黄化衰老,灌浆期叶鞘与上层叶片也迅速黄化衰老[25]. 与早衰突变体psl1,psl3和pse(t)的早衰表现不同,psl4(t)突变体从7叶开始出现叶尖黄化,逐渐向叶中部延伸且不会超过叶中部,叶中部到基部及叶鞘仍然保持绿色,对植株产量影响相对较轻.
水稻衰老过程中涉及的相关基因数量较多,调控路径复杂,具有较为庞大的调控网络[18]. 根据基因的作用途径可分为以下几类:叶绿体的发育及叶绿素降解途径,蛋白质合成、降解及转运途径,激素调控途径,细胞程序化死亡途径等. 各途径既独立又协同,共同调控叶片衰老过程的有序进行. 水稻的NYC1和NOL与叶绿素的降解相关,NYC1,NOL基因编码叶短链脱氢酶/还原酶,均定位在类囊体膜上,在促进叶片衰老过程的叶绿素b降解中发挥重要作用[26]. 弱势早衰突变体wls5的叶绿体发育异常,由于积累了过量的活性氧,导致叶片中的叶绿素严重降解,内源植物激素紊乱[27]. psl4(t)突变体叶片中的叶绿素发生大量降解,但与nyc1,nol和wls5不同的是其叶绿体的类囊体结构也发生降解,且其叶绿体中的淀粉积累量减少. 目前报道的与叶片早衰相关的淀粉积累突变体较少,OsSRT1 RNAi植株在两叶期表现早衰,成熟种子中的淀粉粒变小而松散,且OsSRT1编码水稻中的组蛋白去乙酰化酶,参与调控淀粉在植株中的积累[28]. 叶片早衰淀粉积累异常突变体esl9在叶片中淀粉合成增加,但是淀粉分解速率不及合成速率,导致叶片中合成的淀粉仍不能完全降解,故引起叶绿体中淀粉颗粒积累压迫甚至损伤类囊体[14]. 与esl9不同,psl4(t)叶片中淀粉积累数量明显减少,但淀粉颗粒明显增大,推测可能是淀粉颗粒的积累导致叶绿体中类囊体结构的损伤降解. 遗传分析发现,psl4(t)突变体的早衰性状受单隐性核基因控制;利用分子标记将目标基因定位于第4染色体长臂端两个SSR标记(RM17004和RM17006)之间38.5 kb的范围内,该区间有7个注释基因. 本研究发现叶早衰基因的时空表达与其余衰老基因有一定的差异,说明引起植物叶衰老的遗传基础复杂. 通过对该早衰突变体psl4(t)的遗传分析和基因定位,可以作为叶早衰分子机理研究的互补验证和补充,在水稻的遗传改良和衰老机制解析等方面具有重要意义.
 DownLoad:
DownLoad: