





-
开放科学(资源服务)标识码(OSID):

-
随着全球人口的持续增长,粮食需求日益增加,水稻作为主要粮食作物之一,其产量直接关系到国家的粮食安全。粮食安全不仅是国家发展的基本保障,也是社会稳定的重要基础,因此,提高水稻产量成为水稻育种的核心目标之一。水稻产量主要由单位面积有效穗数、每穗粒数和粒质量3个因素决定,其中粒质量主要受到籽粒形态和灌浆程度的影响。水稻籽粒具有谷类作物种子的典型特征,其内部的胚和胚乳被一层薄种皮包裹,储存灌浆期积累的淀粉和其他营养物质,占据成熟种子的大部分。籽粒外层由内颖和外颖组成的颖壳包裹,在发育早期,颖壳细胞快速分裂以增加细胞数量;而后细胞分裂逐渐减慢,细胞开始扩展以增大面积;最终颖壳不同维度的细胞数目和细胞大小决定了籽粒长度、宽度和厚度,从而限制灌浆充实的程度。因此,籽粒形态主要由颖壳细胞增殖和细胞扩展调节,是与水稻产量密切相关的重要农艺性状[1-2]。
近年来,分子生物学和基因组学的发展为探究水稻粒型的调控机制带来了新的变革。大量调控水稻粒型的数量性状遗传位点(QTL)以及相关基因被成功鉴定与解析[3],其中多数基因通过调控颖壳细胞的增殖或/和扩展来影响水稻粒型,其功能也在随后研究中得到验证。目前,已有多条水稻粒型调控途径被鉴定,如泛素—蛋白酶体降解途径、G蛋白信号途径、植物激素途径以及转录因子调控途径等。这些调控途径的关键基因在水稻籽粒发育进程中担当重要角色,如GRAIN WIDTH 2(GW2)位于泛素—蛋白酶体降解途径,其编码一个RING-like E3泛素连接酶,通过降解其锚定到蛋白酶体的底物负调节细胞分裂。GW2的功能缺失突变体颖壳细胞数目增加,粒宽和粒质量增加[4]。RICE HETEROTRIMERIC G-PROTEIN ALPHA SUBUNIT 1(RGA1)和RICE HETEROTRIMERIC G-PROTEIN BETA SUBUNIT 1(RGB1)基因位于G蛋白信号途径,分别编码G蛋白复合体中的Gα亚基和Gβ亚基,通过正向调控细胞增殖来影响籽粒大小,其功能缺失突变体表现为籽粒变小[5-7]。DWARF 2(D2)和DWARF 11(D11) 位于油菜素内酯(BR)信号途径,编码细胞色素P450家族蛋白,通过正向调控颖壳细胞扩展影响籽粒大小[8-10]。BIG GRAIN 1(BG1)位于生长素(Auxin)信号途径,编码一个膜定位蛋白,通过正向调控颖壳细胞增殖和扩展影响籽粒大小[11]。SHORT AND WIDEN GRAIN 1(SWG1)编码一个转录因子,通过正向调控细胞扩展影响籽粒长度,同时可负向调控横向细胞增殖与扩展影响籽粒宽度,从而在不同方向上呈现出相反的调控效应[12]。GRAIN WIDTH 8(GW8)编码一个包含SBP结构域的转录因子OsSPL16,GW8的高表达可以促进细胞增殖,导致籽粒变宽。GRAIN WIDTH 7(GW7) 编码一个TONNEAU1募集基序蛋白,其表达量上调能增加颖壳纵向细胞分裂并减少横向细胞分裂。GW8能直接与GW7的启动子结合并抑制其表达,最终实现对水稻粒宽的正向调控[13-15]。
此外,丝裂原活化蛋白激酶(MAPK)级联反应也对调控水稻籽粒形态起到了重要作用。MAPK级联反应是真核生物中保守的信号转导机制,由MAPK、MAPK激酶(MKK)和MAPKK激酶(MKKK) 3类激酶通过顺序磷酸化,将外源或发育信号传递至下游靶分子[16-18]。在典型机制中,MKKK在质膜受体刺激下通过磷酸化MKK的丝氨酸/苏氨酸残基使其激活,随后被激活的MKK双重磷酸化MAPK的T-X-Y保守基序中的苏氨酸和酪氨酸残基,最终通过底物的磷酸化实现信号传递[19]。研究表明,植物可利用编码的MKKK、MKK和MAPK蛋白组合成数千种MKKK-MKK-MAPK级联反应,进而调节不同生物学过程[20]。在水稻中OsMKKK10-OsMKK4-OsMAPK6级联反应途径可影响穗部与籽粒形态的发育。SMALL GRAIN 2(SMG2)、SMALL GRAIN 1(SMG1)、DWARF AND SMALL GRAIN1(DSG1) 分别编码OsMKKK10、OsMKK4、OsMAPK6,这3个基因突变均导致植株矮化、穗型直立密集,并因限制颖壳细胞增殖而使籽粒变小。OsMKKK10能与OsMKK4互作并将其磷酸化,OsMKK4继而磷酸化OsMAPK6,形成一条正向调控颖壳细胞增殖的级联反应[21]。此外,GRAIN SIZE AND NUMBER 1(GSN1) 基因编码一个丝裂原活化蛋白激酶磷酸酶OsMKP1,可直接与OsMAPK6互作,并通过去磷酸化使后者失活。当GSN1的表达水平降低时,籽粒虽增大但籽粒数减少;反之,表达水平升高使籽粒数增多但籽粒变小。由此可见,GSN1通过负调控OsMKKK10-OsMKK4-OsMAPK6级联途径,在水稻每穗粒数与籽粒大小之间进行平衡[22]。另外,研究还发现编码WRKY转录因子的OsWRKY53是该级联反应的直接下游靶基因。oswrky53突变体籽粒变小,而超量表达OsWRKY53则使籽粒增大[23-24]。
综上所述,影响籽粒形态的基因数量庞大,且粒型调控网络相当复杂。部分基因上下游的调控关系及作用机制尚不完善,我们仍需要鉴定并克隆更多基因以完善粒型调控网络。本研究中,我们发现了1个与水稻籽粒发育相关的突变体,其主要表现为细胞增殖与扩展异常导致籽粒变小。遗传分析表明,该突变性状受1个隐性单基因控制,我们将其命名为small grain 5 (smg5)。利用MutMap方法,我们将SMG5的候选基因定位在水稻第7条染色体上27.5~29.2 Mb中。结合重测序和PCR测序结果发现,smg5突变体中LOC_Os07g47950基因第5外显子的第37位发生了C-T单碱基替换,从而导致原本翻译的氨基酸由苏氨酸变为异亮氨酸。LOC_Os07g47950基因编码一个丝氨酸/苏氨酸蛋白激酶,其功能此前未被鉴定。本研究为解析SMG5基因调控籽粒形态发育的分子机制,进而完善水稻粒型调控网络奠定基础。
HTML
-
本研究中的水稻小籽粒突变体small grain 5 (smg5)来源于西南大学水稻研究所籼稻保持系西大1B(XD1B)的甲基磺酸乙酯(EMS)诱变库,经过多代自交进行分离纯化后,突变体性状得到稳定遗传。本研究中所用到的水稻材料均种植在重庆市歇马实验基地。
-
在野生型和smg5突变体成熟期选取籽粒,用扫描电镜和体视镜进行表型观察并拍照,抽穗期选取未开花的小穗,置于FAA固定液(由50%无水乙醇,0.9 mol/L的冰乙酸和3.7%甲醛混合配制)中,浸泡超过48 h以固定细胞形态。样本依次进行乙醇梯度脱水、二甲苯透明处理、石蜡浸泡和包埋,调整蜡块形状并固定在石蜡切片机上,切出10 μm厚蜡带;用毛笔轻挑蜡带平放于载玻片上,进行预检查和分段后,转移至涂有多聚赖氨酸的载玻片上,分别于展片机和烘箱中(42 ℃)进行展片与烘片;最后进行番红染色,并用中性树脂封片,用光学显微镜(NikonE600)观察制作好的石蜡切片并拍照。
-
统计分析于2023年夏季水稻抽穗期与成熟期进行。在农艺性状的统计分析中,随机选取10株野生型和10株突变体,统计穗长、一次枝梗数、二次枝梗数、每穗粒数、结实率、粒长、粒宽和千粒质量等农艺性状。在细胞形态和数目统计分析中,随机选取野生型和突变体成熟期的10个籽粒进行扫描电镜观察,统计颖壳纵向细胞数量与细胞长度;随机选取抽穗期10个小穗进行石蜡切片观察,每张切片统计小穗颖壳横向细胞数量与细胞面积。差异显著性均采用t检验,p<0.05表示差异有统计学意义。
-
以西大1B和smg5突变体为亲本进行杂交获得F1代,F1代自交获得F2代。观察并统计F2代群体中野生型表型和突变体表型单株数,计算F2代群体性状分离比并进行卡方检验。
-
基于混合群体分离分析法(Bulked segregant analysis,BSA),将西大1B和smg5突变体杂交,所获F2代群体用于目标基因的定位。F2代群体中出现明显性状分离,在分离群体中选取野生型单株和突变型单株各30株,混取等量叶片分别构建野生型基因池和突变型基因池,将样品送至深圳华大基因股份有限公司进行基因组重测序分析。
将野生型基因池和突变型基因池中的测序结果分别与日本晴参考基因组序列进行比对,运用Sugihara等[25]在GitHub平台上发布的MutMap v2.3.8软件包筛选SNP。首先,运用Trimmomatic软件处理原始测序获得的reads并去除低质量序列;随后,利用BWA和SAMtools软件进行reads比对和初步变异检测,借助BCFtools软件,依据映射质量不低于26、碱基质量不小于18的标准筛选InDel/SNP,计算InDel/SNP指数,去除突变型基因池中与野生型相同的InDel/SNP;最后,用Mutplot软件绘制InDel/SNP在水稻12条染色体上的滑动窗口分布图,据此确定候选基因的定位区间。选取定位区间内在突变型基因池中SNP指数大于0.9的SNP为候选SNP,利用IGV(Integrative Genomics Viewer)浏览器查看候选SNP在基因组上的位置,结合在国家水稻数据中心(
https://www.ricedata.cn/gene/ )查询获得的基因注释信息及表达情况进一步筛选候选基因。 -
从Gramene网站查询并下载候选基因序列,用Vector NTI 10.5软件设计特异性扩增引物,引物名称LOC_Os07g47950,正向序列(5′-3′)GCAGCAGTGTTGTACTGTTCTTCG,反向序列(5′-3′)GGGAGGGCTTACTCTGTATCAGG,分别以野生型、smg5突变体以及F2群体中正常表型植株和突变表型植株的DNA为模板扩增目标序列。PCR总体系为25.0 μL:含10× PCR buffer 12.5 μL,dNTPs 5 mmol/L,ddH2O 4.4 μL,10 μmol/L的前后引物各0.8 μL,模板DNA 1.0 μL,5 U/μL Taq酶0.5 μL。PCR程序为:94 ℃预变性5 min;94 ℃变性30 s,57 ℃退火30 s,72 ℃复性1 min,35个循环;72 ℃延伸5 min。PCR扩增产物经1%琼脂糖凝胶电泳后,送至北京擎科生物科技有限公司进行测序分析,利用Vector NTI 10.5软件对测序结果进行序列比对,用Chromas2软件查看测序峰图,确定突变位点并完成候选基因鉴定。
1.1. 试验材料
1.2. 形态与组织学分析
1.3. 统计分析
1.4. 遗传分析
1.5. 基因定位
1.6. 候选基因分析与克隆
-
与野生型(WT)相比,smg5突变体成熟期植株的株型与穗型发生了明显变化。相较于野生型,smg5突变体植株矮化,表现为株型整体紧凑;突变体穗型直立,而野生型植株穗多且下垂(图 1a、1b)。对穗部各项农艺性状进行统计分析,发现突变体穗长平均为26 cm,略长于野生型的25.5 cm(图 1c);一次枝梗数平均为14.2个,较野生型显著增加了22.99%(图 1d);二次枝梗数平均为24.7个,较野生型减少了25.5%(图 1e);此外,突变体平均每穗颖花数和每穗粒数分别为145个和50.8个,较野生型分别降低14.51%和71.6%(图 1f、1g),后者的大幅降低导致其结实率极显著低于野生型,仅为34.6%(图 1h)。由此推断,smg5突变体的穗型变化可能与其二次枝梗数和每穗粒数的降低有关。
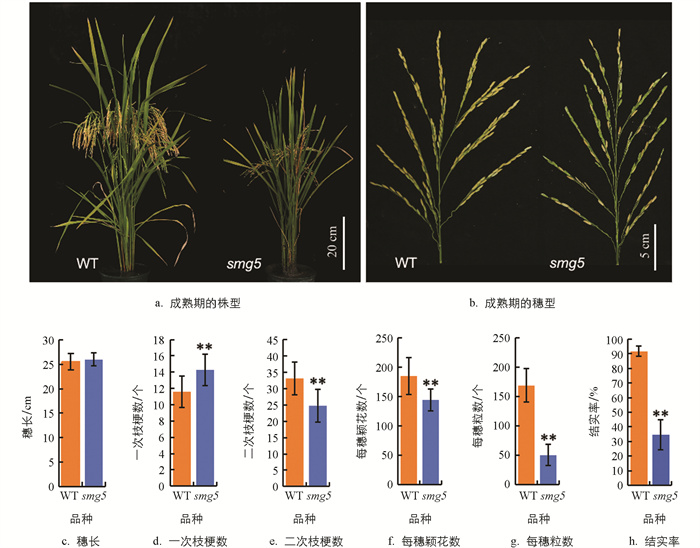
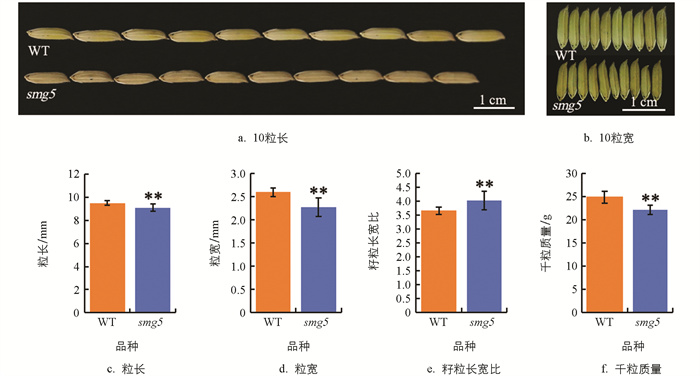
smg5突变体与野生型(WT)在籽粒形态上也存在显著差异。与野生型相比,smg5突变体籽粒显著变小,其粒长降低了4.28%,平均为9.1 mm(图 2a、2c);粒宽降低了12.6%,平均为2.22 mm(图 2b、2d);长宽比为4.03,相较于野生型增加了11.1%(图 2e),整体呈现小籽粒表型。由于smg5突变体粒长、粒宽均降低,其千粒质量比野生型降低了10.69%,平均为22.22 g(图 2f)。
-
水稻籽粒形态主要由颖壳细胞的大小与数目决定,为进一步探究smg5突变体籽粒变小的原因,我们对野生型和突变体成熟籽粒的颖壳进行了组织细胞学分析。利用扫描电镜对野生型和突变体籽粒颖壳的纵向细胞特征进行观察,结果显示,二者内外稃外表皮细胞均发生明显硅化,并在每个细胞上分布有瘤状突起,细胞间分布有不均一毛刺(图 3a、3e)。统计籽粒内外稃交界处纵向排列的细胞总数,发现突变体相较于野生型减少了7.43%(图 3i)。统计籽粒内外稃交界处相同部位2 mm范围内的细胞个数,发现二者均约为23个,表明籽粒外稃纵向排列细胞长度差异无统计学意义(图 3b、3f、3j)。因此,突变体籽粒长度降低主要是外稃纵向排列细胞增殖水平降低、细胞数目减少所致。
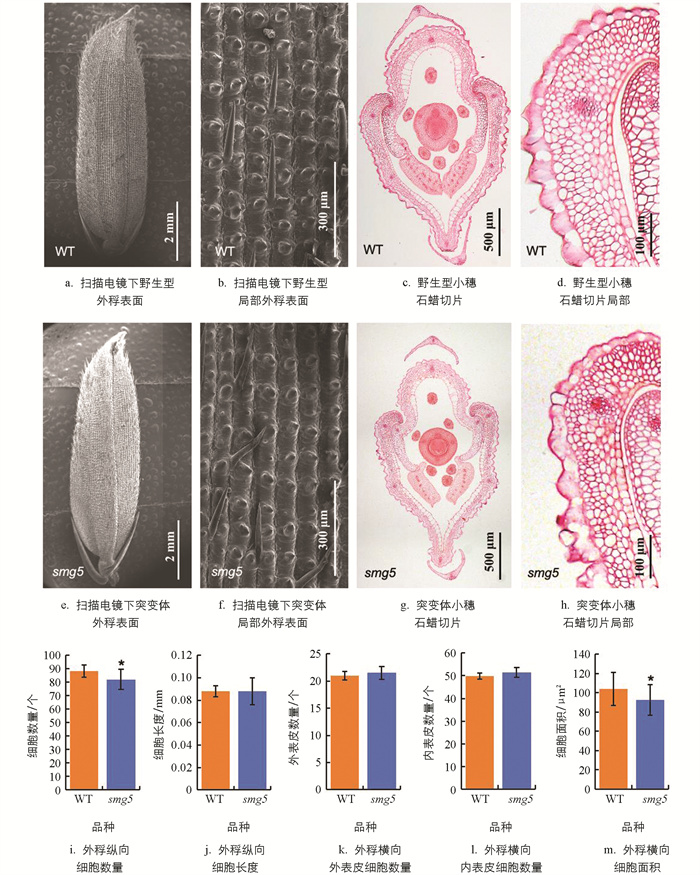
通过石蜡切片,进一步分析和统计野生型和突变体开花期小穗颖壳的横向细胞特征。观察发现,野生型和突变体内外稃细胞组成和特征无明显差异,由外向内依次由1层明显硅化的外表皮细胞、2~5层染色较深且面积较小的厚壁细胞、2层染色较浅且面积较大的薄壁细胞和1层大小不一的内表皮细胞组成(图 3c、3g)。统计突变体与野生型外稃横向外表皮层的细胞数量,发现二者均约为21个(图 3k);统计突变体与野生型外稃横向内表皮层细胞的数量,发现二者均约为50个(图 3l)。结果表明,突变体与野生型籽粒外稃细胞的横向增殖能力差异无统计学意义。由于颖壳横截面横向排列细胞在线性排列上无明显规律,无法准确测量细胞的平均宽度,故统计外稃相同位置的细胞数目与面积,计算发现突变体外稃横向细胞的面积平均为92 μm2,显著低于野生型(图 3d、3h、3m),表明突变体外稃细胞的横向扩展能力较低。综上表明,突变体籽粒变窄主要是外稃横向细胞扩展水平降低、细胞面积减小所致。SMG5基因可通过正向调控颖壳纵向细胞增殖与横向细胞扩展,分别影响籽粒长度和宽度。
-
smg5突变体与XD1B杂交产生的F1代植株表现正常,F2代植株表现出显著的性状分离,其中,正常植株143株,smg5突变表型植株57株,总和为200株,分离比为2.509∶1。经卡方检验其分离比符合3∶1(χ2=1.307<χ0.052=3.84),表明smg5突变体的性状受1对隐性单基因控制。
-
以smg5突变体与野生型XD1B为亲本杂交后的F2代为定位群体,基于BSA法,构建了目标性状的分组基因池:30株野生型构成野生型基因池、30株突变体构成突变型基因池。对分组基因池进行重测序后,分别从野生型基因池和突变型基因池获得73 709 928个和78 564 001个reads,在水稻基因组的覆盖率分别为98.56%和98.49%。使用MutMap软件筛选获得2 314个InDel和2 082个SNP,计算获得其在两个基因池中的InDel/SNP指数(I)。
基于遗传学原理进一步筛选InDel/SNP:①突变型基因池单株表型较准确,引起突变表型的位点在基因池单株中应大部分发生突变,理论上I≈1。由于测序过程中可能产生其他错误导致出现假阳性位点,规定I≥0.9;②野生型基因池包含野生型单株(即引起突变表型的位点均未发生突变,I=0)以及杂合突变单株(即引起突变表型的位点在同源染色体中的1条染色体发生突变,I=0.5),规定I=0~50%。以1 500 kb为窗口大小,150 kb为步长,分别绘制InDel/SNP指数在水稻12条染色体上的滑动窗口分布图(图 4a,4b)。InDel指数除在第7条染色体末端有较明显起峰外,于12条染色体间分布差异不大(图 4a);但SNP指数在第7条染色体末端有明显峰点,表明该区域上SNP与突变表型紧密连锁(图 4b)。由此将候选基因定位于水稻第7条染色体末端,其突变由SNP变异导致。以1 000 kb为窗口大小,100 kb为步长,绘制SNP指数在第7条染色体上的滑动窗口分布图(图 4c)。选取突变型基因池中SNP指数大于等于0.9的12个SNP为候选SNP,其在野生型基因池中的SNP指数均符合0~50%的理论比例。由图 4c可知,候选SNP中有7个集中分布于第7条染色体上27.5~29.2 Mb中。
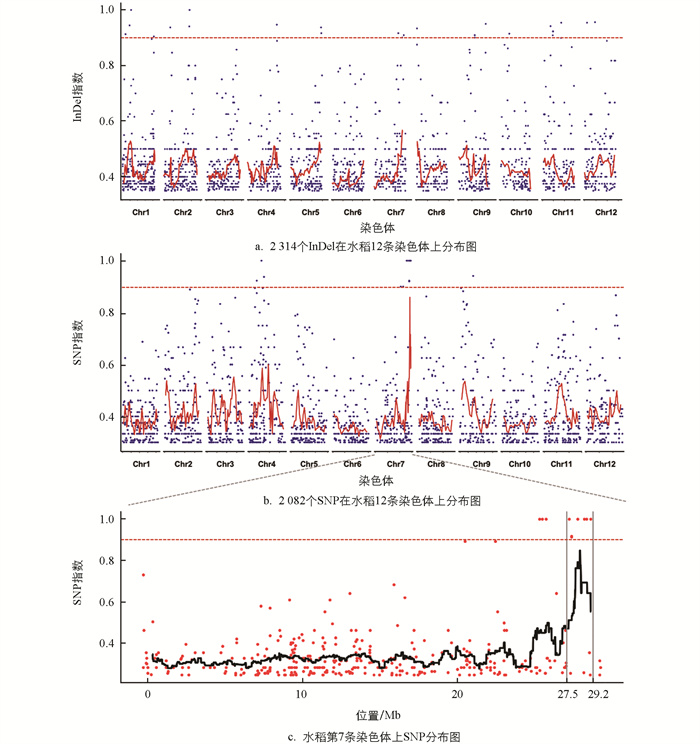
利用IGV软件查看候选SNP在水稻基因组中的位置,在国家水稻数据中心网站上查询对应基因在水稻中的注释信息及表达情况。由表 1可知,有6个SNP位于基因上,仅有3个基因(LOC_Os07g38130、LOC_Os07g43270、LOC_Os07g47950)的SNP位于外显子区域,其余SNP位于内含子区域。结合注释信息进一步分析SNP的突变效应,发现LOC_Os07g38130基因编码一个多聚半乳糖醛酸酶抑制蛋白,其上SNP突变为苏氨酸的无义突变;LOC_Os07g43270基因编码一个SKP1蛋白,其上SNP突变为丙氨酸的无义突变;LOC_Os07g47950基因编码一个丝氨酸/苏氨酸蛋白激酶,其上SNP突变代表苏氨酸变为异亮氨酸的有义突变。LOC_Os07g47950在水稻花穗中表达量较高,与籽粒发育过程中颖壳形态的建成相适应。综合以上结果,我们将LOC_Os07g47950基因暂定为SMG5的候选基因。
-
为进一步确定候选基因,比对LOC_Os07g47950基因上SNP在BSA混池中的差异,并利用PCR测序进一步验证。结果显示,该基因上SNP在突变型基因池、野生型基因池和野生型亲本中的SNP指数分别为1.0、0.2和0,符合遗传学比例。PCR扩增结果显示,6株smg5突变体的LOC_Os07g47950基因第5个外显子的第37位均发生了C-T单碱基替换,导致原本翻译的氨基酸由苏氨酸(Thr)变为了异亮氨酸(Ile)(图 5a)。通过NCBI网站分析LOC_Os07g47950基因及其蛋白信息,发现LOC_Os07g47950基因位于水稻第7条染色体长臂端,包含1 563个碱基。其编码一个包含521个氨基酸的丝氨酸/苏氨酸蛋白激酶,其中第193到458位为Ser/Thr蛋白激酶结构域(S_TKc)。smg5突变体中苏氨酸—异亮氨酸的突变恰好位于该结构域中(图 5b)。
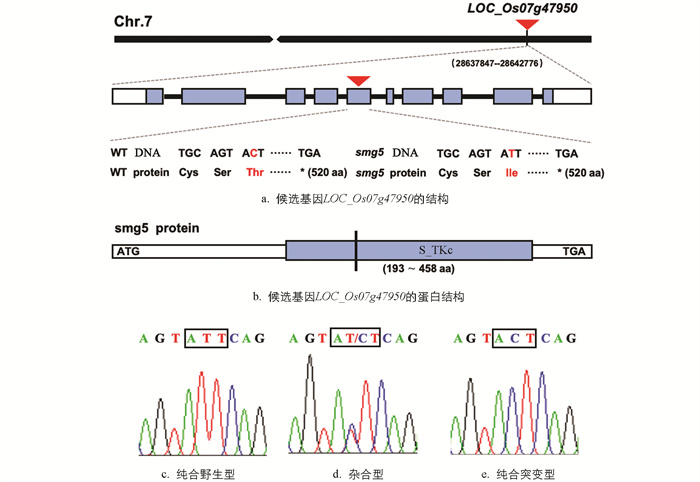
从定位群体F2中随机选择10株野生型植株和8株突变体植株的叶片提取DNA,利用PCR测序进一步验证,SNP分型统计结果见表 2。Chromas软件分析显示,smg5突变体亲本中LOC_Os07g47950基因为纯合突变型,突变率为100%;10株正常表型的植株为纯合野生型(图 5c)或杂合型(图 5d),突变率为15%;8株突变表型植株为纯合突变型(图 5e),突变率为100%。以上结果表明,smg5突变体的小籽粒表型确认与LOC_Os07g47950中的C-T碱基突变有关。
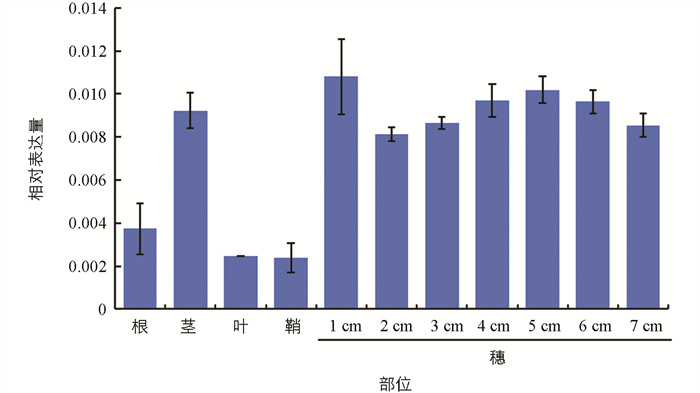
运用qRT-PCR对候选基因LOC_Os07g47950进行表达模式分析,结果显示,该基因在水稻中组成型表达,尤在穗中表达量较高(图 6),这与网站查询所得的表达情况相符。综合以上结果,我们最终将LOC_Os07g47950基因定为SMG5的候选基因。
2.1. smg5突变体的表型分析
2.2. smg5突变体颖壳的组织细胞学分析
2.3. 遗传分析
2.4. SMG5基因定位
2.5. SMG5候选基因鉴定
-
水稻籽粒形态主要由颖壳细胞增殖和细胞扩展调节。近年来,已鉴定到许多与籽粒发育相关的基因,其中已知多个调控小籽粒形态的基因,他们主要通过单独调控细胞增殖使籽粒变小。例如,SMG2/OsMKKK10正向调控颖壳细胞增殖,smg2突变体外稃纵向细胞和横向细胞的数量均显著低于野生型,致使其粒长和粒宽减少[26];SMG1/OsMKK4相应突变体与野生型相比,其颖壳表皮细胞面积差异不显著,但细胞数量显著降低,说明SMG1/OsMKK4主要影响细胞增殖调控粒型[27];DSG1/OsMAPK6正向调控籽粒外稃表皮细胞增殖,其突变体粒长与粒宽均降低[28]。此外,还有基因通过单独调控细胞扩展影响籽粒形态。例如,small grain 1(smg11) 突变体籽粒的颖壳外表皮细胞面积显著减小,但细胞数量与野生型相似,说明SMG11主要影响颖壳细胞扩展正向调控籽粒大小[29]。目前所鉴定的小籽粒表型相关基因,大多数通过单独影响细胞增殖或细胞扩展调控籽粒形态,但能同时调控细胞增殖和扩展使籽粒变小的基因较为少见。
本研究中,通过组织细胞学分析深入解析了smg5突变体籽粒变小的原因。在纵向上,突变体外稃纵向细胞长度与野生型差异无统计学意义,但纵向细胞总数显著低于野生型,说明SMG5基因通过调控细胞增殖影响籽粒长度;在横向上,突变体籽粒颖壳外稃的细胞数目与野生型相似,但相同部位单个细胞面积显著低于野生型,表明SMG5基因通过影响细胞扩展调控籽粒宽度,因此,SMG5基因能够同时正向调控纵向细胞增殖和横向细胞扩展,从而影响籽粒的形态发育。深入探究其粒型调控机制有益于解析这一特殊的调控方式。
-
MutMap是Abe等[30]于2012年提出的一种基因克隆方法。基于BSA法,MutMap结合高通量二代测序技术,通过比对突变型与野生型基因池的测序结果及等位基因频率,降低遗传背景的干扰,准确定位到与目标性状紧密连锁的InDel/SNP突变位点上[31]。随后,Sugihara等[25]于2022年在GitHub平台上发布MutMap v2.3.8软件包,在Abe等的基础上优化算法,使测序数据处理速度提升了5~6倍,并可以精确识别出基因池间的InDel/SNP差异。与图位克隆等传统的基因克隆方法相比,MutMap无需构建复杂的遗传群体和遗传图谱。基于全基因组测序数据,使得该方法能够更全面、准确、迅速地锁定目标基因。MutMap方法综合运用了全基因组学、生物信息学和分子遗传学等多学科技术,在不断实践与优化中成为一种成熟的基因克隆方法,为水稻等作物的育种研究提供了丰富的基因资源。
MutMap方法现已被广泛应用于克隆水稻的各项产量性状基因,并在籽粒发育相关基因研究中展现出较大潜力,如调控小籽粒形态的基因SMG2/OsMKKK10、SMG11/D2等,均通过MutMap方法进行了克隆。SMG2/OsMKKK10编码蛋白激酶参与MAPK途径调控籽粒形态[26],SMG11/D2编码细胞色素P450参与BR植物激素途径影响籽粒形态[29]。此外,王涛[32]运用MutMap方法克隆到长籽粒基因GRAIN LENGTH 5(GL5),其编码一个酪氨酸磷酸酶蛋白。GL5可与MAPK6互作,参与协调MAPK级联反应调控粒型,因此MutMap对于鉴定水稻粒型新基因有重要意义。
本研究运用MutMap v2.3.8软件包筛选并绘制InDel/SNP指数在水稻12条染色体上的滑动窗口分布图,结合遗传学原理,将发生SNP变异的SMG5候选基因定位于水稻第7条染色体上27.5~29.2 Mb内,根据基因信息最终将候选基因暂定为LOC_Os07g47950,该基因编码一个丝氨酸(Ser)/苏氨酸(Thr)蛋白激酶,其功能尚未被鉴定。
结合PCR测序结果可知,smg5突变体中LOC_Os07g47950基因发生的C-T单碱基变化导致苏氨酸—异亮氨酸变化,此突变恰好位于该基因所编码的Ser/Thr蛋白激酶结构域中。在籽粒大小调控OsMKKK10-OsMKK4-OsMAPK6级联反应途径中,OsMKK4激酶和OsMKKK10激酶也同样含有Ser/Thr蛋白激酶结构域。OsMKKK10可与OsMKK4相互作用并磷酸化OsMKK4,但MKKK10中Ser/Thr蛋白激酶结构域中的点突变会导致其对MKK4的磷酸化活性消失[21]。另有研究表明Ser/Thr残基的保守性磷酸化通常对于MAPK活化十分关键,活化的MAPKKKs首先磷酸化MKKs的Ser/Thr残基,随后活化的MKKs对下游MAPKs保守结构域进行双重磷酸化,从而触发MAPKs对下游底物的活化[33]。候选基因LOC_Os07g47950中单碱基突变所导致的氨基酸变化可能会改变其所编码的激酶活性,从而影响其对上下游激酶的磷酸化/去磷酸化过程,最终调控籽粒形态,但其具体的分子机制仍需后续试验进一步验证。综上,克隆SMG5基因,通过功能分析解析SMG5调控粒型的分子机制,对于完善水稻粒型调控网络具有重要意义。
-
本研究从籼稻保持系XD1B的EMS诱变库中鉴定到1个水稻小籽粒突变体smg5。与野生型相比,smg5突变体粒长、粒宽和千粒质量显著降低;植株整体紧凑且矮化,穗型直立,且二次枝梗数、每穗颖花数、每穗粒数和结实率均显著低于野生型。组织细胞学分析表明,SMG5基因通过正向调控颖壳纵向细胞增殖和横向细胞扩展,分别影响籽粒的长度和宽度。遗传分析表明,该突变体性状受1对隐性单基因控制。利用XD1B和smg5突变体构建F2分离群体,采用MutMap方法将LOC_Os07g47950基因暂定为SMG5的候选基因。PCR测序显示,LOC_Os07g47950基因的第5个外显子的第37位发生了C-T单碱基替换,导致原本编码的氨基酸由苏氨酸变为了异亮氨酸。该突变位点恰好位于其所编码的Ser/Thr蛋白激酶结构域中。本研究进一步丰富了与籽粒形态相关的突变体材料,为深入理解水稻粒形调控网络提供了基因资源。
 DownLoad:
DownLoad: