


 下载:
下载:
-
开放科学(资源服务)标志码(OSID):

-
柑橘是我国最重要的水果,主要种植在南方各省市的山地和丘陵地区. 柑橘产区横跨几个气候带,许多柑橘产区的降雨量季节分配不均匀,因此季节性缺水造成的干旱是制约我国许多地区柑橘产业发展的一个主要因素[1]. 植物有多种抗旱和耐旱机制应对干旱胁迫,以维持正常的生长和发育. 近年的研究发现,一些小分子RNA不仅与植物的生长发育密切相关,而且在植物的抗旱中也发挥着至关重要的作用[2]. 小分子RNA主要有3种类型,siRNAs,miRNAs和piRNAs,其中miRNAs是目前生物体内存在最广泛的,也是研究最多的一类小RNA. 这些小RNA长度大约在20~24个核苷酸,它们通过碱基互补配对与靶基因结合,进而在转录后水平调控基因表达,促使mRNA降解或抑制其翻译,起负调控靶基因的作用[3-4]. miRNA通过调控靶基因参与植物的生长发育和逆境响应[5],许多研究报道指出,有多种miRNA参与植物的干旱胁迫响应. miR196g是最早在水稻中被发现的与干旱相关的miRNA[6],豇豆中发现了有近20个miRNA以不同的方式响应干旱胁迫[7],干旱逆境诱导碧桃叶片及根部中262和368个miRNA差异表达[8],miR168和miR396在拟南芥和烟草中的表达也受干旱胁迫诱导[9]. 在模式植物拟南芥中,miR171在干旱、低温、低氧、低磷、低硫等胁迫下表达量都会发生变化[10]. 豆科植物中miR171也响应干旱胁迫[11]. 在水稻中,过表达osa-miR171f可以通过调控类黄酮生物合成基因的表达来增强水稻的耐旱性[12],它通过调节SCL6-I和SCL6-II的转录水平在抗旱性中发挥作用. 在苹果中,敲除mdm-miR171i和过表达它的靶基因MsSCL26.1均可增强其抗旱性[13]. 进一步研究发现,mdm-miR171通过调节抗氧化基因表达和抗坏血酸代谢来响应干旱胁迫. 以上研究结果表明,miR171家族成员响应植物的干旱胁迫. 目前的研究表明,miR171家族的功能高度保守. 因此可以推测柑橘中的Cre-miR171也可能响应干旱胁迫,但柑橘中Cre-miR171是否响应干旱胁迫以及调控的靶基因的研究还未见报道.
为明确Cre-miR171对柑橘耐旱的作用,本研究通过对Cre-miR171在不同柑橘砧木品种、组织及根系不同部位、发育时期的表达特征进行分析,对Cre-miR171过表达的柑橘砧木枳和模式植物拟南芥进行耐旱性及根系形态评价,探究Cre-miR171对植物响应干旱胁迫的影响. 并在转基因柑橘中对预测的靶基因进行表达分析,为深入研究Cre-miR171调控柑橘耐旱性的分子机理提供参考.
全文HTML
-
柑橘砧木品种枳(Poncirus trifoliata Raf.)、扁平橘(Citrus depressa Hayata. ‘Shiikuwasha’)、红柠檬(C. limonia Osbeck. ‘Canton Lemon’)的种子采自国家果树种质柑橘圃(重庆,北碚). 本氏烟(Nicotiana benthamiana)、拟南芥(Arabidopsis thaliana)种子为本实验室自繁.
DNA提取试剂盒(植物基因组DNA快速提取试剂盒),反转录试剂盒(Revert Aid First Strand cDNA Synthesis Kit),RNA提取试剂盒(天根DP504),限制性内切酶(Thermo Fisher scientific),DNA Marker (Takara),引物合成和测序由北京擎科生物科技有限公司完成.
-
将枳、扁平橘和红柠檬的种子去除种皮后于(28±1) ℃下保湿催芽. 挑选萌芽一致的种子播种于膨胀珍珠岩∶蛭石(体积比1∶1)的基质中,在温室(25±1) ℃中培养. 选取一月龄左右的不同砧木幼苗,取其根、茎、叶用于Cre-miR171的组织表达分析. 分别于枳种子播种后30 d,60 d和90 d,取全根用于根的不同发育时期Cre-miR171的表达分析. 于枳种子播种后45 d,取根尖和侧根发生区样品用于Cre-miR171在根不同部位的表达分析. 于枳种子播种后60 d,取茎、叶、侧根发生区以及根尖用于Cre-miR171靶基因的表达分析. 以上取样各5株,设5个生物学重复.
选取长势一致的1月龄枳的水培幼苗,分别用15%的PEG6000及100 μM的脱落酸(ABA)水溶液进行胁迫处理,在处理的0 h,3 h,6 h,12 h,24 h分别采集全根作为样品,用于胁迫处理下Cre-miR171的表达分析. 选取播种后2个月、长势一致的盆栽枳幼苗于温室内进行干旱处理,在干旱处理的0 d,5 d,10 d,15 d分别采集根尖区与侧根发生区的组织样品,用于干旱胁迫下Cre-miR171的表达分析.
-
Cre-miR171的前体序列来自本实验室前期构建的小RNA文库. 利用Tbtools对Cre-miR171前体序列与从柑橘基因组数据库下载得到的克里曼丁橘(C. clementina)基因组数据进行序列比对,获得Cre-miR171前体序列上游3 000 bp启动子序列,其顺式作用元件通过PlantCARE(
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )进行预测分析. 利用psRNATarget:A Plant Small RNA Target Analysis Server (2017 Update) 在线网站psRNATarget(2017 update)软件进行小RNA靶基因的互补配对,以miRNA的5′端为种子序列与参考基因组克里曼丁橘的转录本3'UTR区对Cre-miR171的靶基因进行预测. 靶基因序列从在线网站Phytozome(https://phytozome-next.jgi.doe.gov/pz/portal.html )上获取. -
采用Primer 3在线软件设计1对Cre-miR171前体序列的特异引物,分别在序列的两端加上SwaI,BamHI酶切位点(表 1),以枳的DNA为模板,对Cre-miR171进行克隆. 用Solution I连接酶将Cre-miR171前体序列的全长与经SwaI,BamHI双酶切后的PGBi载体进行连接,转化到大肠杆菌DH5a感受态细胞中,挑选单克隆送往北京擎科生物科技有限公司进行测序. 经测序验证正确的过表达载体转入农杆菌EHA105感受态细胞,用枳上胚轴进行柑橘的遗传转化[14],采用花序侵染法进行拟南芥的遗传转化[15].
-
利用PCR方法对Cre-miR171的过表达柑橘植株和拟南芥植株进行鉴定. 正向引物是35S启动子一段序列,反向引物为Cre-miR171前体序列(表 1),以转基因植株的DNA为模板进行扩增. 采用茎环法qRT-PCR检测各转基因柑橘植株的Cre-miR171相对表达量,选择Cre-miR171表达量最高的3株进行嫁接扩繁. 同时对其靶基因的表达进行定量分析.
-
构建两类烟草瞬时表达载体进行Cre-miR171与其靶基因的互作验证. 一类为Cre-miR171前体序列超表达的GUS载体,将Cre-miR171的前体序列与线性化的GUS载体连接;另一类为改造过的超表达载体pMS4载体,pMS4载体在GFP基因前后有两个酶切位点XhoI和XbaI,经双酶切之后用于插入miRNA靶基因位点. 靶基因位点是通过人工合成的两条寡核苷酸链退火后形成的双链,分为S Cclscl6,SCclscl22,SCclscl26以及经过改造的MTSCclscl6,MTSCclscl22,MTSCclscl26两类(表 2). 重组后的质粒经北京擎科生物科技有限公司测序验证正确后,转化至农杆菌GV3101菌株的感受态细胞,在烟草中利用农杆菌介导的侵染共同表达miRNA及其靶基因.
-
茎环qRT-PCR和qRT-PCR分别被用来分析miRNA及其靶基因的相对表达量. miRNA的内参基因为U6,靶基因的内参基因为柑橘CsActin. 用Primer5软件设计miRNA的反转录引物和靶基因、内参基因的实时荧光定量PCR引物(表 3). qRT-PCR反应体系为:2×NovoStartⒸ SYBR qPCR SuperMix 5 μL,正、反向引物各0.2 μL,RNase Free Water 2.6 μL,cDNA 2 μL(500 ng). 扩增程序为:95 ℃预变性10 min,然后95 ℃ 15 s,58 ℃ 60 s,共设置40个循环. 所有的定量反应都在实时PCR System (Applied Biosystems 7 500 Fast,Applied Biosystems,USA)上完成. 每个qRT-PCR反应重复3次. miRNA及其靶基因的相对表达量采用2-ΔΔCT法进行计算[16].
-
选择长势一致的过表达Cre-miR171的拟南芥植株进行26 d的干旱处理,并观察其在干旱情况下的表型变化. 在处理14 d时取叶片样,测定POD与脯氨酸的含量.
选择长势一致的枳野生型和转基因植株,剪取相同生长部位的叶片,放置在干燥的滤纸上,测定脱水处理60 min内的质量变化,计算相对失水率. 对脱水处理后的叶片进行电导率的测定,计算处理后叶片的相对电导率.
剪取对照组和Cre-miR171表达量最高的过表达枳的枝条,扦插于蛭石基质中,待其长出根系后,评价分析根的生长发育表现.
1.1. 试验材料
1.2. 试验方法
1.3. 生物信息学分析
1.4. 过表达Cre-miR171载体的构建及遗传转化
1.5. 转基因植株的鉴定与其靶基因的筛选
1.6. 烟草瞬时表达载体的构建与转化
1.7. Cre-miR171及其靶基因的荧光定量分析
1.8. 转基因植株的耐旱性评价
-
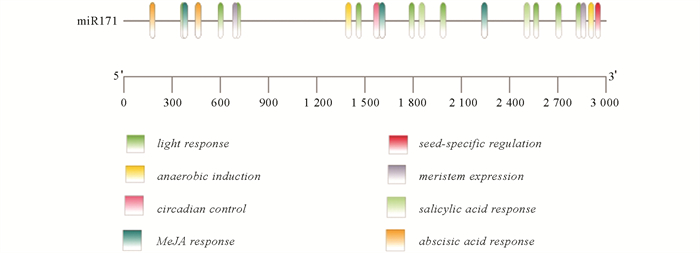
利用PlantCARE对Cre-miR171前体上游启动子顺式作用元件进行预测(图 1),结果发现:该小RNA的启动子区域除了含有典型的具有转录起始功能的TATA-box,CAAT-box核心元件外,还包括激素和胁迫响应相关元件,如脱落酸、水杨酸和茉莉酸甲酯响应元件. 此外,还包含一些光响应、厌氧、昼夜响应等相关元件. 根据以上结果,推测Cre-miR171的表达受多种因素的诱导,能参与逆境胁迫下的应答及调控.
-
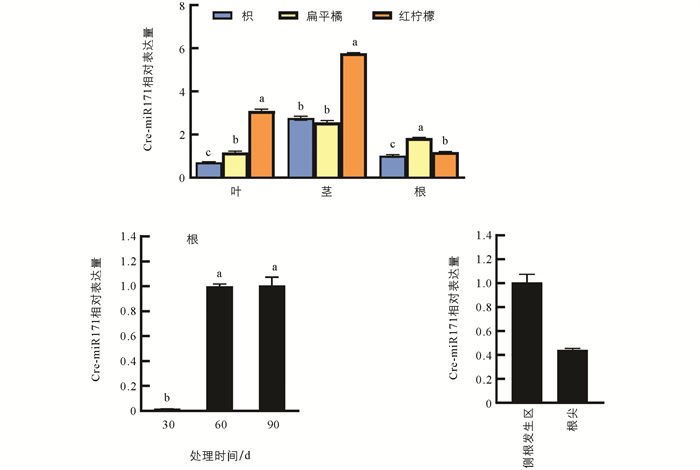
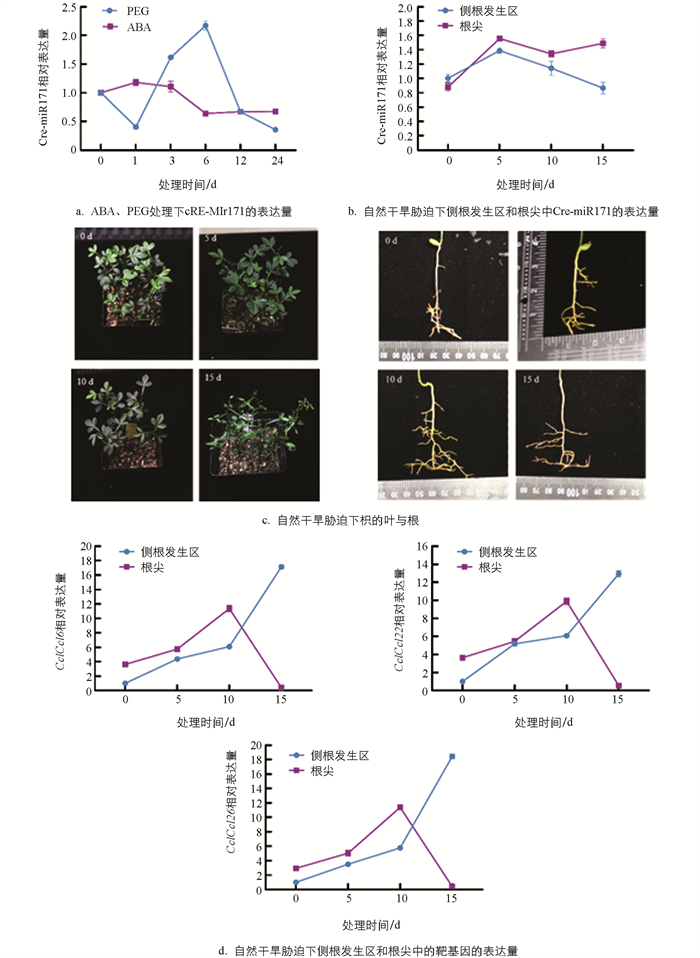
在枳、扁平橘、红柠檬的幼苗中对Cre-miR171的组织表达模式进行分析,这3种砧木的耐旱性明显不同,其中枳的耐旱性最强[15]. Cre-miR171在3种不同砧木中表现出相同的组织表达特征,在根中的表达量相对较低,在茎中的表达量最高. 同时,Cre-miR171在耐旱性较强的枳的根和叶中的表达量显著低于其他2种耐旱性比较弱的砧木,与砧木的耐旱性相反,说明Cre-miR171可能参与柑橘对干旱胁迫的响应. Cre-miR171在枳根生长发育的不同时期的表达也存在明显差异,Cre-miR171在种子发芽后第30 d时表达极低,在30~60 d时急剧上升,在60~90 d时表达量无明显变化,表明Cre-miR171可能与根系的早期发育相关. Cre-miR171在根系的表达有一定的组织特异性,在侧根发生区的表达量明显高于根尖(图 2),说明Cre-miR171对侧根发生和根伸长生长的作用存在差异.
-
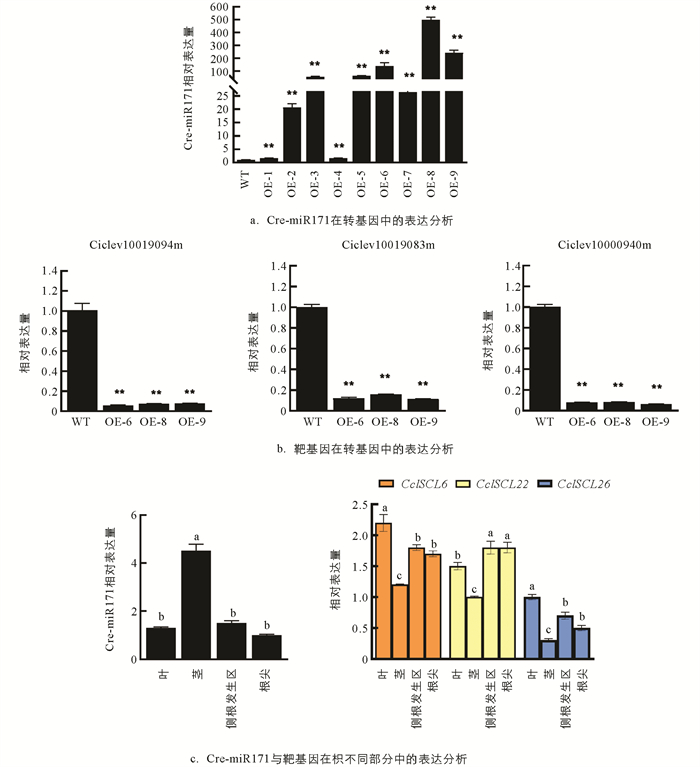
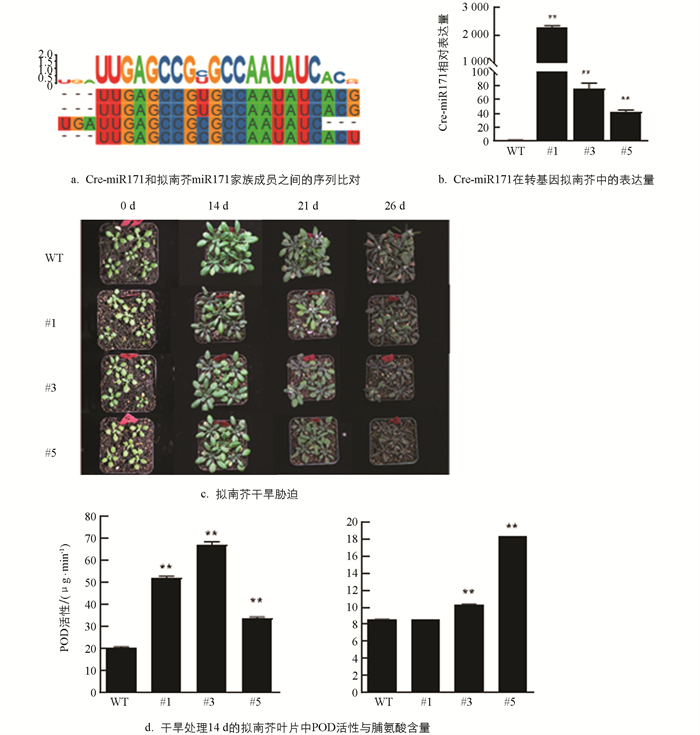
利用psRNATarget对Cre-miR171的靶基因进行了预测,通过生物信息学分析去除大量重复基因后,发现Cre-miR171主要靶向9个靶基因,分别为SCL家族基因Ciclev10019094m(SCL6),Ciclev10019083m(SCL22),Ciclev10000940m(SCL26);GRAS基因Ciclev10018916m、酰基活化酶相关基因Ciclev10003943m、蛋白磷酸酶相关基因Ciclev10001232m、乙烯不敏感基因Ciclev10000617m以及2个功能未知基因Ciclev10010118m,Ciclev10027698m. 在嫁接成活的9株超表达Cre-miR171的枳植株中选取Cre-miR171表达量上调最高的OE-6,OE-8,OE-9等3个株系对靶基因的表达进行定量分析(图 3a),发现Ciclev10019094m,Ciclev10019083m和Ciclev10000940m等3个靶基因的表达在超表达植株中受到了明显的抑制,说明这3个靶基因明显受Cre-miR171的负调控,靶向关系较强(图 3b). 基因注释和同源基因比较分析指出,这3个基因是同属于SCL家族的同源基因,在本研究中将这3个基因分别命名为CclSCL6,CclSCL22,CclSCL26. 这3个靶基因在枳中的表达表现出相同的组织特异性(图 3c),和Cre-miR171的表达均呈明显负相关,进一步验证了这3个基因是Cre-miR171的靶基因,在功能上有明显的相似性.
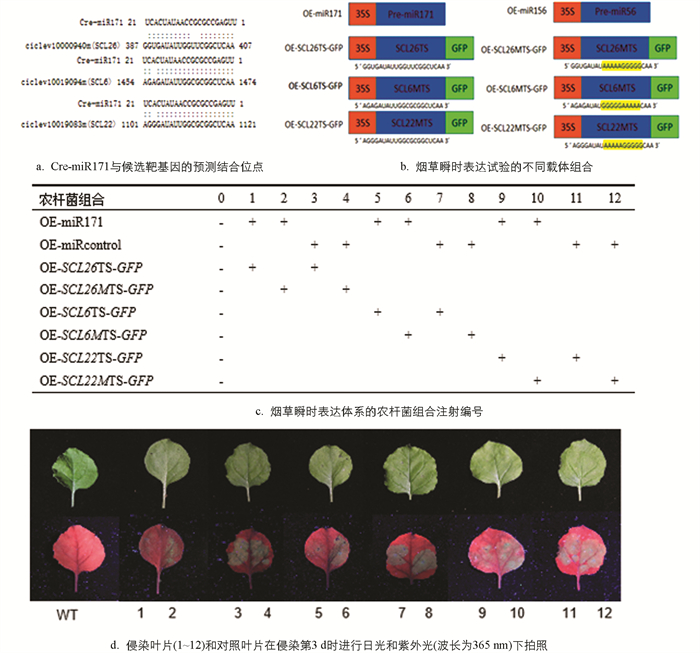
利用烟草瞬时表达试验对Cre-miR171与CclSCL6,CclSCL22,CclSCL26的相互作用进行了验证(图 4). 用农杆菌侵染烟草叶片,分别共同表达Cre-miR171、Cre-miRcontrol、携带靶位点的GFP基因和携带修饰后的靶位点的GFP基因(图 4b). 结果发现,含有OE-miR171和OE- SCL26-GFP,OE- SCL6-GFP,OE- SCL22-GFP的农杆菌共同侵染烟草叶片后,叶片中GFP荧光信号均发生了明显的减弱,而其他组合叶片中的GFP荧光信号没有发生明显的改变(图 4d). 因此表明在植物体内,Cre-miR171能够抑制靶基因CclSCL26,CclSCL6,CclSCL22的表达.
-
ABA处理下,Cre-miR171的表达在根中呈现出逐渐下降的趋势,但是变化幅度较小;PEG6000处理对Cre-miR171的表达影响非常显著,处理过程中,Cre-miR171在根中的表达呈现出先下降后快速上升再快速下降的趋势,表现出对胁迫处理的强烈响应,说明Cre-miR171对渗透胁迫比较敏感(图 5a). 为了了解干旱胁迫下Cre-miR171在根的不同部位的表达,对枳幼苗进行了为期15 d的自然干旱处理. 在干旱处理的第10 d时,观察到侧根开始发生(图 5c),根尖数目增加,而地上部分形态变化不明显. Cre-miR171的表达明显响应了干旱胁迫,其在根尖中的表达量高于侧根发生区,特别在干旱处理10 d后更为显著(图 5b),3个靶基因的表达也显示出了对干旱胁迫的明显响应,并在处理10 d后出现剧烈的变化(图 5d),与Cre-miR171在这期间的表达明显负相关. 从整个处理时期看,Cre-miR171与靶基因在干旱胁迫下根的两个部位的表达均没有达到显著的相关性(表 4),特别在根尖的表达相关性更弱. 但在处理的第5 d后,Cre-miR171与其靶基因在根的两个部位的表达趋势均转变为明显的负相关(表 5),特别在10 d后均表现出强烈的变化,说明Cre-miR171的表达受干旱胁迫的诱导,并在响应胁迫后对靶基因产生了较强的调控作用. 另外,Cre-miR171在根部两个部位的表达在处理的第10 d后出现相反的趋势,3个靶基因的表达也随之发生相应的变化,说明Cre-miR171在干旱胁迫下能抑制根的伸长生长,促进侧根的发生.
-
miR171在植物中较为保守[17],序列比对显示,Cre-miR171与拟南芥miR171家族成员之间只有2个核苷酸的差异(图 6a). 对过表达Cre-miR171 3周龄的拟南芥进行26 d的干旱处理,结果显示,在干旱14 d内,野生型拟南芥的生长状况良好,叶片色泽鲜绿,而转基因植株的叶片颜色则明显加深;在干旱21 d时转基因拟南芥叶片的边缘枯黄,野生型的拟南芥叶片的颜色也开始加深,但转基因拟南芥的叶片颜色更深,而且转基因拟南芥植株的生长已受到了极大的抑制,植株明显矮小;干旱26 d时转基因拟南芥的叶片因脱水而导致叶片大面积萎蔫(图 6c),干旱胁迫处理对转基因株系伤害程度明显高于野生型株系. 以上结果说明,过表达Cre-miR171使拟南芥对干旱逆境的敏感性显著增加. 干旱处理14 d时转基因植株的过氧化物酶含量高于野生型植株,其脯氨酸含量也显著高于野生型(图 6d),也说明了转基因拟南芥更早地响应干旱胁迫.
-
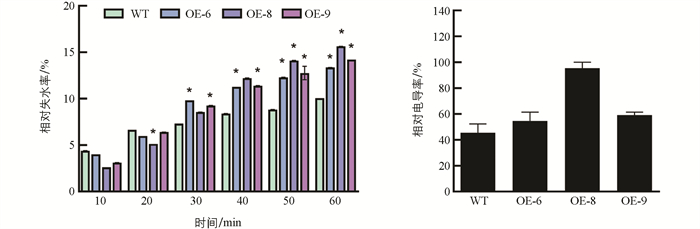
对扩繁成活的转基因植株OE-6,OE-8,OE-9的叶片进行脱水处理,在处理30 min到60 min时转基因株系叶片的相对失水率均显著高于野生型. 转基因枳的叶片在脱水处理后的电导率高于野生型(图 7),说明转基因枳膜系统被干旱损伤的程度高于野生型,超表达Cre-miR171明显降低了枳的耐旱能力.
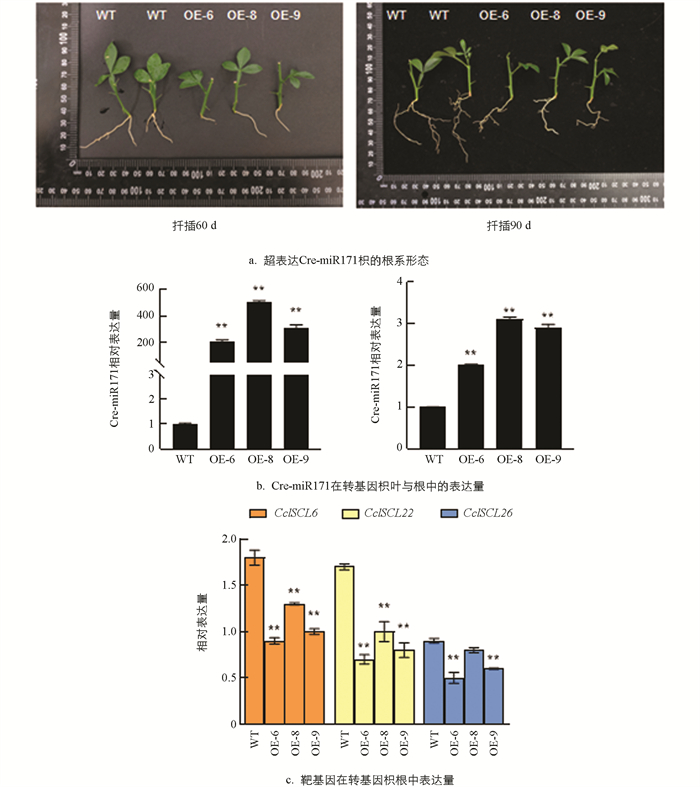
对扦插生根的对照组和超表达Cre-miR171的枳植株的根系进行形态观测(图 8a),发现转基因植株的侧根比对照组的少,根系也明显短,说明Cre-miR171影响了枳根系的生长发育,超表达Cre-miR171抑制了枳的主根伸长生长和侧根发生. 对扦插90 d的转基因枳进行表达定量分析,结果显示,Cre-miR171在转基因枳的叶片与根中的表达量都显著上调(图 8b),其靶基因CclSCL26,CclSCL6,CclSCL22的表达量受到抑制(图 8c),说明Cre-miR171可能通过负调控其靶基因CclSCL26,CclSCL6,CclSCL22的表达来抑制主根的伸长生长和侧根的发生.
2.1. Cre-miR171前体上游调控区相关启动子元件分析
2.2. Cre-miR171在柑橘中的表达特征
2.3. Cre-miR171靶基因的预测、鉴定和表达分析
2.4. 非生物胁迫下Cre-miR171与靶基因在根中的表达分析
2.5. 表达外源Cre-miR171拟南芥的耐旱性评价
2.6. 超表达Cre-miR171枳的耐旱性评价
-
在植物中,miR171参与调节植物基本的发育和代谢过程[18-19]. 在模式植物拟南芥中,miR171通过调控其靶基因AtSCL6-II,AtSCL6-III和AtSCL6-IV的表达影响茎尖分生组织[20-21]. 过表达miR171前体可以抑制腋芽形成和花芽数量,从而改变拟南芥茎的结构[22]. 拟南芥幼苗在高盐、低温和干旱胁迫下miR171的表达量都会上调[9],推测miR171参与了拟南芥非生物胁迫的调控. 番茄中miR171和SCL6通过调节激素信号去调控番茄的生长发育[23]. 甘蔗中miR171响应盐胁迫[24].
Cre-miR171启动子区有光信号、脱落酸、水杨酸、茉莉酸甲酯等激素和胁迫响应元件,同时miR171被证实响应光信号[25]、茉莉酸甲酯[26]、水杨酸、脱落酸[27]诱导,表明Cre-miR171可能受光信号、茉莉酸甲酯、水杨酸、脱落酸等多条信号通路的影响,推测Cre-miR171可能参与逆境胁迫下的应答及调控. 表达Cre-miR171的拟南芥对干旱胁迫更敏感,脱水处理对过表达Cre-miR171枳叶片的膜系统的伤害显著高于对照,说明过表达Cre-miR171的枳转基因植株的耐旱能力降低. 这与在苹果中的研究结果相似[13],表明Cre-miR171对柑橘的耐旱性有负调控的作用.
有研究表明miR171在根的发育和生长中发挥重要作用,miR171参与苹果[28]和葡萄[29]的不定根形成,miR171还影响了水稻的根系发育[30],同时miR171被证实可以影响蒺藜苜蓿的侧根密度[31]. 本研究中,Cre-miR171在柑橘根发育的早期高表达,而且在侧根发生区的表达量高于根尖,表明Cre-miR171对柑橘根系的发育及侧根的形成有重要影响.
本研究中预测并验证的3个Cre-miR171的靶基因(CclSCL6,CclSCL22,CclSCL26)是植物中特有的转录因子,其功能具有多样性,参与植物的生长发育[32]、信号转导、生物胁迫和非生物胁迫相关的应答过程[33]. Cre-miR171在响应干旱胁迫后在侧根发生区下调表达,在根尖上调表达,趋势相反,3个靶基因的表达也随之发生相应的显著变化,说明在干旱胁迫下,Cre-miR171能通过对靶基因表达的调控抑制根的伸长生长,促进侧根的发生. 可以推测,Cre-miR171可能调控柑橘根系在干旱胁迫下的可塑性.