


 下载:
下载:
-
开放科学(资源服务)标识码(OSID):

-
水稻是我国最重要的粮食作物之一, 提高其产量和品质对于保障我国粮食安全、增进民生福祉具有极为重要的意义[1]。随着粮食缺口的逐步扩大和生物技术的不断发展, 传统的杂交选育方法已经很难满足水稻生产的实际需要, 种质创新成为亟待解决的重要课题。近年来, CRISPR/Cas9等基因编辑技术的出现, 为水稻等作物的遗传改良提供了新的工具[2]。利用基因编辑技术创新种质, 通过对选定的基因进行特定修饰, 可以有针对性地引入理想性状, 实现作物的改良[3-8]。Gn1a(Grain number 1a)是水稻第1染色体短臂上的主效QTL, 通过调控细胞分裂素(Cytokininin, CK)代谢影响每穗粒数, 在高产育种中具有重要的应用价值[6-7, 9]。Gn1a编码的OsCKX2蛋白由FAD结合结构域和CKX催化结构域组成, 其中FAD结合结构域负责结合FAD(黄素腺嘌呤二核苷酸), 作为电子受体介导细胞分裂素侧链的氧化反应, 而CKX催化结构域是酶的催化核心, 两者协同作用, 特异性降解细胞分裂素, 从而调控水稻的生长发育[4, 6]。Gn1a表达量的降低可导致花序分生组织中细胞分裂素的积累, 促进一次、二次枝梗的分化, 最终提升每穗粒数和产量[9-10]。Gn1a是水稻产量性状改良的重要基因编辑靶点, 重穗型恢复系蜀恢498Gn1a基因第3外显子上11 bp碱基缺失会引起CKX催化结构域编码序列的移码突变, 使蛋白失活并显著增加每穗粒数[7]。此外, 在籼稻中花11的遗传背景下, 将Gn1a基因序列前端的第135~157位碱基敲除, 也能引起类似效应[6, 10]。一方面, 在转基因水稻中利用RNA干扰(shRNA)抑制Gn1a表达, 可以显著增加分蘖数和穗粒数并提升水稻的产量[11]; 另一方面, OsMPK6与锌指转录因子DST的相互作用可以激活Gn1a的转录, 使水稻的每穗粒数显著减少[12]。这些研究表明, 通过Gn1a关键位点的精确编辑可用于改良水稻的产量性状。当前Gn1a研究主要集中在CKX催化结构域, 而FAD结合结构域的功能解析仍较少, 限制了该位点在种质创新中的应用[9-16]。本研究利用CRISPR/Cas9技术在籼稻渝金香(YJXWT)中Gn1a基因的第1外显子上FAD结合结构域的编码序列进行靶向敲除, 筛选、鉴定出不含有外源基因的纯合单株, 成功创制出穗粒数增加、具有实际应用价值的突变体, 并系统分析其农艺性状、基因表达模式及单倍型变异, 为进一步筛选适宜的基因编辑位点、优化水稻高产育种策略提供理论依据和种质资源。
全文HTML
-
籼稻品种渝金香(YJXWT)由西南大学水稻研究所提供, 种植于歇马实验基地, 所有材料常规管理。Cas9载体PC1300-UBI-Cas9[14]由王克剑老师提供。大肠杆菌菌株(Escherichia coli)DH5α和农杆菌菌株(Agrobacterium tumefaciens)LBA4404由武汉艾迪晶生物科技有限公司(
http://www.edgene.com.cn/ )保存。 -
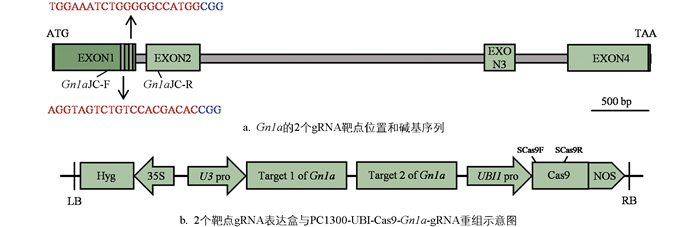
利用CRISPR/Cas9技术进行定向基因编辑, 依据Gn1a (LOC_Os01g10110)基因序列特征, 利用CRISPR-GE网站(
http://skl.scau.edu.cn/ )设计靶点[17], 在第1外显子区选择583~585 bp处的GGC作为靶点1的PAM序列, 586~605 bp处20 bp作为靶点1; 660~662 bp处的GGC作为靶点2的PAM序列, 663~682 bp处20 bp作为靶点2(图 1a)。参照刘建国等[18]的方法, 在正反向引物5′端分别添加接头GGCA和AAAC(表 1), 引物双链化经AarI酶进行酶切后的PC1300-UBI-Cas9载体进行连接获得重组植物表达载体PC1300-UBI-Cas9-Gn1a-gRNA(图 1b)。通过农杆菌介导法转化籼稻品种渝金香(YJXWT)的成熟胚愈伤组织, 并通过潮霉素抗性筛选获得阳性转基因单株。遗传转化、潮霉素抗性筛选步骤均由武汉艾迪晶生物科技有限公司(http://www.edgene.com.cn/ )完成, 获得的水稻组培苗种植于田间用于后续试验。 -
利用The Gramene Database植物基因组学数据库(
https://www.gramene.org/ )鉴定和分析本研究中基因编辑靶点在籼稻全基因组序列中的特异性, 导入双靶点序列, 结合The Gramene Database中现有的籼稻基因组序列进行分析。综合以上分析结果进一步确定双靶点是否在全基因组中具有特异性。 -
为了检测水稻基因敲除株系中Gn1a突变的具体类型, 针对包含双靶点所在区域附近500 bp的碱基序列设计了Gn1aJC-F和Gn1aJC-R引物(图 1a、表 1), 引物由北京擎科生物科技股份有限公司重庆分公司(
https://www.tsingke.com.cn/ )合成。使用CTAB法[4]提取野生型水稻渝金香(YJXWT)和基因敲除单株水稻植株的叶片DNA作为模板进行PCR扩增, 通过分析扩增产物的片段长度确定基因的不同突变类型; 进一步将扩增产物送至北京擎科生物科技股份有限公司重庆分公司进行测序, 以YJXWT的DNA测序结果为对照, 分析编辑植株中Gn1a基因的突变情况。 -
检测基因敲除成功的水稻单株在基因编辑过程中T-DNA的插入情况, 针对转基因载体上800 bp左右的碱基序列设计SCas9F和SCas9R引物(图 1b、表 1), 由北京擎科生物科技股份有限公司重庆分公司合成。利用基因敲除单株的叶片DNA为模板, 同步设置阴、阳对照进行PCR扩增, 通过观察琼脂糖凝胶电泳的条带判断T-DNA的插入情况。
-
提取野生型水稻渝金香(YJXWT)的DNA进行测序, 获得完整的Gn1a基因编码区序列。使用NCBI(
https://www.ncbi.nlm.nih.gov/ )平台上的ORFFinder工具(https://www.ncbi.nlm.nih.gov/orffinder/ )进行开放阅读框(ORF)的查找及翻译; 将Gn1a基因的核苷酸序列粘贴至输入框, 选择合适的遗传密码表翻译得到可能的蛋白质序列; 选择最长且符合生物学意义的开放阅读框对应的蛋白质序列用于后续结构域预测。进入NCBI的CDD搜索页面(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ), 在搜索框中粘贴上述操作获得Gn1a基因对应的蛋白质序列, 启动结构域预测分析。 -
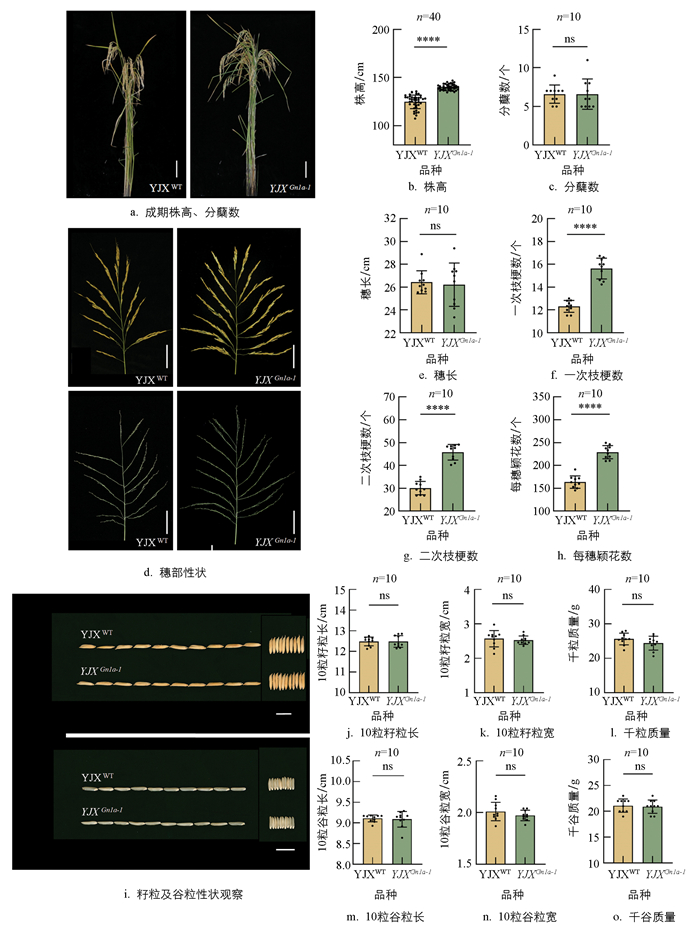
经测序鉴定后将基因编辑成功的植株幼苗单本移栽到歇马实验基地中, 依据常规稻田管理方式进行栽培。待水稻成熟时, 选择野生型水稻渝金香(YJXWT)和Gn1a编辑植株, 使用单反相机(CANON EOS 70D)进行拍摄, 收集表型数据。在成熟野生型水稻渝金香(YJXWT)和YJXGn1a-1突变体中选取40株长势基本一致的植株考察株高, 选取10株长势基本一致的植株考察分蘖数、穗长、一次枝梗数、二次枝梗数、每穗颖花数、粒长、粒宽、千粒质量、单株总小穗数、单株总籽粒数、结实率和单株产量等田间农艺性状, 采用t检验进行统计分析。
-
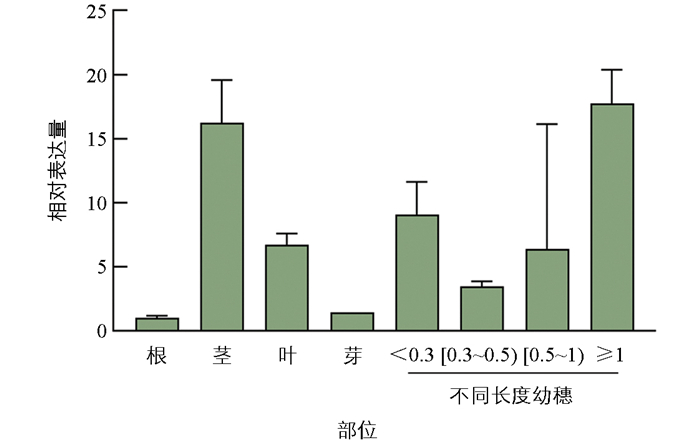
在孕穗期, 分别取常规籼稻品种渝金香(YJYWT)的根、茎、叶和不同长度的幼穗于液氮中速冻, 提取RNA进行反转录合成cDNA用于实时荧光定量PCR。利用荧光定量PCR的预混体系和Bio-RadX96 PCR仪, 以Actin作为内参(表 1), 设3次重复, PCR扩增程序为: 95 ℃预变性60 s; 95 ℃变性15 s; 60 ℃退火15 s; 70 ℃延伸60 s, 共40个循环。所得数据使用2-ΔΔCt法计算相对定量值(RQ)与误差值, 结合数据分析Gn1a在植株不同部位的表达模式。
-
利用Rice SNP-Seek Database(
https://snp-seek.irri.org/ )、Rice Variation Map v2.0(http://ricevarmap.ncpgr.cn/ ) 鉴定和分析单核苷酸多态性(single nucleotide polymorphisms, SNP)及插入/删除(Insertion/Deletion, InDel)位点, 结合Rice SNP-Seek Database中现有的水稻表型数据进行单倍型分析[19-20]。综合以上分析结果进一步对目标性状和SNP、InDel标记位点进行关联性分析。
1.1. 试验材料
1.2. CRISPR/Cas9靶点设计及PC1300-UBI-Cas9基因编辑载体构建
1.3. CRISPR/Cas9靶点特异性分析
1.4. 转基因突变体的检测鉴定
1.5. 编辑植株的T-DNA插入鉴定
1.6. 结构域分析
1.7. YJXGn1a-1突变体的农艺性状考察
1.8. Gn1a表达模式分析
1.9. 单倍型分析
-
在籼稻全基因组序列中对靶点展开特异性分析, 结果显示: 两个靶点的期望值(E-val)均为0.001 4, 说明双靶点在籼稻基因组中无随机匹配, 匹配高度特异(表 2)。
-
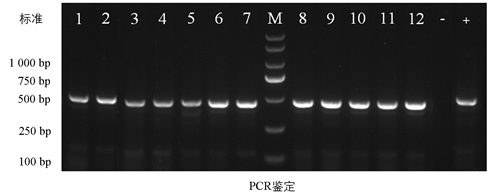
由图 2可知, 从T0代69个单株中提取叶片DNA, 利用PCR扩增和测序分析Gn1a基因第1外显子靶点区段(500 bp), 结果显示, 双靶点设计有效提高了编辑效率和位点突变的准确性。在69个检测单株中, 62株(89.9%)发生了Gn1a缺失突变, 其中28株(45.2%)为纯合缺失突变体, 34株(54.8%)为杂合缺失突变体。
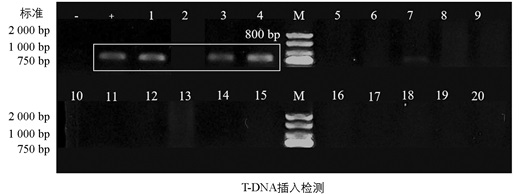
为避免外源载体在T0代植株基因组中的整合导致Cas9蛋白的持续表达, 进一步对单株进行T-DNA插入检测。由于后续农艺性状鉴定所需的单株数为10株, 故随机选择28株纯合缺失突变体中的20株利用PCR扩增检测T-DNA序列, 并通过琼脂糖凝胶电泳分析其插入情况(图 3)。结果显示, 20株纯合缺失突变体中, 仅3株(1、3、4号)检测到T-DNA插入, 其余17株未检测到。17株不含外源T-DNA的突变体可用于后续试验, 并可作为稳定遗传的种质资源加以利用。
-
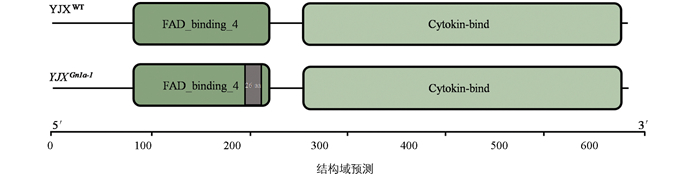
为精确解析编辑靶点的突变情况, 对T0代测序数据进行深入分析, 以确定YJXGn1a-1突变体FAD结合结构域编码区的具体缺失位点。结果显示, Gn1a第1外显子编码区存在78 bp碱基缺失(表 3), 起始于第589位碱基, 终止于666位碱基, 该缺失属于整码缺失突变。
进一步通过蛋白结构域分析, 预测FAD结合结构域的缺失对蛋白功能的影响。结果表明, 在野生型YJXWT中共鉴定出两个功能结构域, 分别为FAD结合结构域(81~238 aa, 共158 aa)和CKX催化结构域(269~581 aa, 共313 aa)。相比野生型, YJXGn1a-1突变体因78 bp碱基缺失, 导致FAD结合结构域内26个氨基酸(197~222 aa)缺失(图 4)。
-
为探究Gn1a蛋白FAD结合结构域缺失对表型的影响, 首先对野生型和YJXGn1a-1突变体的株高及分蘖数进行测量和统计分析。结果显示, 与野生型相比, YJXGn1a-1突变体的株高显著增加, 平均株高由125.0 cm提高至139.8 cm(p<0.01)(图 5a、5b)。分蘖数在突变体与野生型间差异无统计学意义(p>0.05), 均维持在7个左右(图 5a、5c)。YJXGn1a-1突变体的穗长与野生型间差异无统计学意义(p>0.05)(图 5d、5e), 但一次枝梗数和二次枝梗数均显著增加(p<0.01)。突变体平均一次枝梗数为16个, 显著高于野生型的12个(p<0.01); 二次枝梗数为46个, 显著高于野生型的30个(p<0.01)(图 5d、5f、5g)。一次、二次枝梗数的显著增加进一步导致每穗颖花数的大幅提升, 由野生型的164个增加至229个(p<0.01)(图 5d、5h)。
水稻穗粒数增加通常会导致粒型减小[2, 6, 21-22]。为探究YJXGn1a-1突变体穗粒数增加是否影响粒型, 对野生型和突变体的籽粒及糙米性状进行了统计分析。结果显示, 野生型与突变体的10粒籽粒长约12.5 cm, 10粒籽粒宽约2.5 cm, 千粒质量约25 g, 二者差异无统计学意义(p>0.05)(图 5i-5l)。进一步去除谷壳后分析, 10粒谷粒长约9.0 cm, 10粒谷粒宽约2.0 cm, 千谷质量约21 g, 野生型与突变体间差异也无统计学意义(p>0.05)(图 5j、5m-5o)。YJXGn1a-1突变体每穗粒数的显著增加并未引起籽粒性状的显著改变。
-
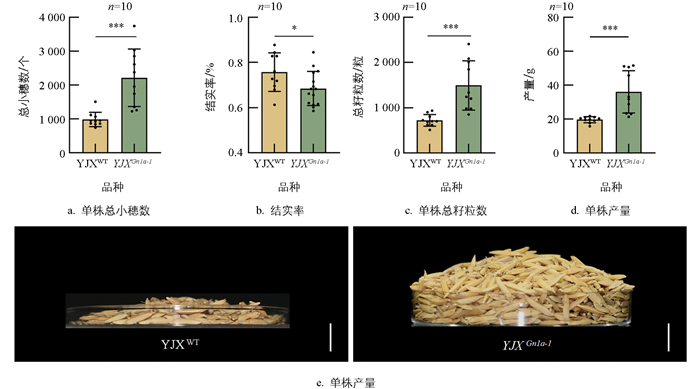
YJXGn1a-1突变体在分蘖数无显著变化的情况下, 一次枝梗数和二次枝梗数均显著增加, 导致单株总小穗数由野生型的989颗极显著增加至2 218颗(p<0.01)(图 6a), 突变体的结实率相较于野生型显著下降(p<0.05)(图 6b), 单株总籽粒数和产量极显著提升(图 6c、6d), 其中单株总籽粒数由725颗增至1 496颗(p<0.01), 单株产量由20 g提高至36 g(p<0.01)(图 6d、6e)。综上表明, Gn1a蛋白FAD结合结构域的缺失可显著提高水稻产量。
-
为探究YJXGn1a-1突变体株高和穗粒数增加的潜在机制, 利用qRT-PCR分析Gn1a在水稻不同组织中的表达模式。结果显示, Gn1a主要在茎及不同长度幼穗中表达(图 7), 表明其可能参与了株高和花序的发育调控。
-
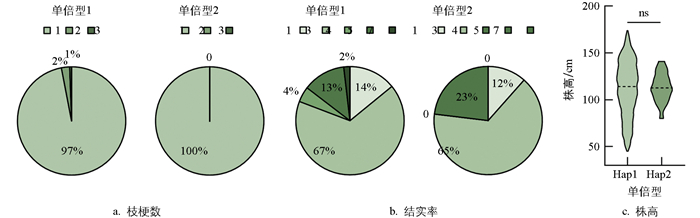
FAD结合结构域编码序列的缺失能够使突变体的株高和单株产量极显著提升。为进一步探究Gn1a缺失区段(Chr1: 5274920-5274997)是否存在具有类似敲除效应的自然单倍型, 利用Rice SNP-Seek Database和Rice Variation Map v2.0对3 024份水稻种质资源进行分析。结果显示, Gn1a基因序列中共鉴定出43个SNP位点和29个InDel位点。在缺失区段内, 仅包含1个SNP位点(Chr1: 5274962), 导致氨基酸由丙氨酸变为苏氨酸, 对应两种不同的单倍型, 且未发现InDel变异。进一步分析该SNP变异对水稻枝梗数、结实率及株高的影响, 发现两种不同单倍型的表现具有一致性, 其中大部分品种枝梗数的量化指标为1, 结实率的量化指标为3, 平均株高约112.5 cm(p>0.05)(图 8)。上述结果表明, 该SNP变异可能对水稻穗粒数和株高的提升无显著贡献, 暗示Gn1a功能的实现可能依赖于其FAD结合结构域的完整性。
2.1. CRISPR/Cas9靶点特异性分析
2.2. FAD结合结构域缺失突变体YJXGn1a-1的鉴定
2.3. YJXGn1a-1突变体Gn1a基因编辑位点分析及结构域变化预测
2.4. FAD结合结构域缺失突变体YJXGn1a-1的农艺性状分析
2.5. FAD结合结构域缺失突变体YJXGn1a-1的产量性状分析
2.6. Gn1a基因在水稻中的表达模式分析
2.7. Gn1a自然单倍型的表型分析
-
水稻产量由单位面积内有效穗数、每穗粒数和粒质量等因子决定, 而这些产量决定因子则是由数量性状位点(Quantitative Trait Locus, QTLs) 控制[17, 23-24]。筛选与穗粒数相关的理想QTL位点有利于培育高产水稻品种[23, 25-27]。Gn1a(OsCKX2)是控制水稻每穗粒数的重要QTL, 编码一种细胞分裂素氧化酶, 其表达下降会导致细胞分裂素在花序分生组织中积累, 从而增加每穗粒数, 提高产量[9-10, 16, 23-24]。Gn1a蛋白由FAD结合结构域和CKX催化结构域组成。CKX是催化活性核心, 而FAD结合结构域则负责结合辅酶黄素腺嘌呤二核苷酸(FAD), 作为电子受体完成细胞分裂素异戊烯基侧链的氧化反应。两个结构域相互协同, 通过β-氧化途径特异性降解细胞分裂素从而调控水稻的生长[9, 11, 15, 28-30]。研究发现, Gn1a基因不仅能够调控穗粒数, 也影响结实率和分蘖数[31]。Wang等[7]研究表明, CKX催化结构域部分缺失可显著增加穗粒数但降低了结实率; Yeh等[11]利用CRISPR/Cas9敲除CKX催化结构域编码区特定位点, 发现其可提高穗粒数但减少了分蘖。本研究利用CRISPR/Cas9基因编辑技术在籼稻品种渝金香(YJXWT) Gn1a基因的第1外显子上设计双靶点敲除, 筛选、鉴定到不含外源基因的YJXGn1a-1突变体。结构域分析表明, YJXGn1a-1突变体存在78 bp碱基缺失, 引起FAD结合结构域中26个氨基酸缺失(图 4)。相较于野生型, YJXGn1a-1突变体的一次枝梗数、二次枝梗数极显著增加, 而籽粒形态和千粒质量并未发生显著变化(图 5), 最终单株产量显著提高(图 6)。值得注意的是, YJXGn1a-1突变体植株高度显著增加。基因的表达分析显示, Gn1a基因在茎及不同长度幼穗中均有表达(图 7), 提示其不仅调控穗粒数, 还可能影响株高, 这一功能或由FAD结合结构域介导。在未来研究中, 可进一步解析Gn1a在株高调控中的作用机制, 并探索利用其编辑效应提高水稻地上生物量, 以创制高光效品种用于育种[32]。
随着CRISPR/Cas9基因编辑技术的发展, 研究者可快速实现目标基因的定向编辑, 以加速水稻的遗传改良[12, 33]。解析高产单倍型突变位点并结合CRISPR/Cas9技术, 可筛选出具有育种价值的突变体[25-26]。已有研究表明, Gn1a基因的缺失是重穗型水稻Shuhui498(R498)每穗粒数增加的决定性因素, 且部分携带InDel突变位点的水稻单倍型表现出显著的穗粒数增加[6-7]。此外, 部分InDel位点与粳籼亚种分化密切相关[33], 可能在水稻高产性状的形成中发挥着关键作用。目前对于水稻InDel位点与表型的关联性分析研究仍停留在不同品种InDel位点的鉴定层面[34-38], 缺乏系统的突变体表型数据收集及数据库构建, 导致育种过程中难以精准筛选关键的InDel位点, 限制了高效创制具有目标性状的水稻种质。本研究基于CRISPR/Cas9技术成功敲除Gn1a蛋白的FAD结合结构域, 并获得YJXGn1a-1突变体, 可用于分析FAD结合结构域缺失的表型效应[39-40]。为了进一步评估编辑位点的育种潜力, 我们在自然种质资源中筛选了编辑区段(Chr1: 5274920-5274997)携带SNP和InDel的单倍型, 用以评估其表型效力。在敲除区间内, 仅检测到1个SNP位点Chr1: 5274962, 其编码氨基酸发生了改变(由丙氨酸变为苏氨酸), 对应两种单倍型。数据库分析显示, 这两种单倍型在株高和枝梗数方面差异无统计学意义(图 8), 表明单一SNP位点的变异不足以作为水稻穗粒数提升的决定性因素, 而FAD结合结构域的完整性对Gn1a功能行使具有重大影响。相较之下, 本研究创制出的YJXGn1a-1突变体表现出株高和每穗颖花数的显著提升, 因此InDel突变及大片段缺失可能比单点突变对Gn1a功能的影响更为深远。未来可针对InDel相关单倍型进行系统的表型收集, 构建InDel位点与水稻高产表型的关联性数据库, 以筛选Gn1a基因与表型强关联的关键InDel位点。结合自然种质资源的InDel突变体筛选及CRISPR/Cas9技术, 可创制具有优良特性的高产单倍型, 以加速水稻品种的改良。
-
表达模式分析表明, Gn1a在水稻茎和不同长度幼穗中特异表达, 提示其可能在水稻株高和花序发育中发挥重要作用。结构域预测表明, Gn1a蛋白由FAD结合结构域和CKX催化结构域组成。本研究以籼稻品种渝金香(YJXWT)为受体材料, 利用CRISPR/Cas9技术靶向敲除Gn1a基因FAD结合结构域编码区段的78 bp碱基序列, 测序检测出无外源片段插入的纯合突变体YJXGn1a-1。农艺性状调查显示, 突变体植株株高显著增加, 一次、二次枝梗数显著增加, 每穗颖花数显著提升, 单株产量显著提高。单倍型分析进一步验证了FAD结合结构域的缺失可显著增加水稻株高和穗粒数。本研究初步解析了FAD结合结构域缺失对株高和穗粒数的影响, 为Gn1a基因的功能解析提供了新的视角, 创制了可用于高产育种的新种质资源, 为培育高产水稻品种提供了理论依据和实践支持。